
第一章 鳥類的演化
1.1 什麼是鳥類?
鳥類是一群獨特而多樣的脊椎動物,屬於恐龍類的後裔。它們是唯一存活至今的恐龍後代,在演化過程中發展出許多特殊的適應性特徵。鳥類最顯著的特徵是羽毛的存在,這是它們與其他動物最明顯的區別。羽毛不僅提供了飛行的能力,還具有保溫、防水、偽裝和求偶等多種功能。
鳥類的身體結構高度適應飛行生活。它們擁有中空的骨骼,這使得骨骼既輕巧又堅固。鳥類的前肢演化成翅膀,與強大的胸肌一起支持飛行。大多數鳥類都具有飛行能力,但也有一些物種,如鴕鳥和企鵝,已經失去了這種能力,轉而適應其他生活方式。
鳥類的呼吸系統非常高效,包括肺部和氣囊系統,能夠在飛行時提供充足的氧氣。它們的心臟相對體型來說很大,可以支援高強度的飛行活動。鳥類的消化系統也經過特殊適應,能夠快速處理食物,減輕飛行時的負擔。
在感官方面,鳥類通常具有極佳的視力,許多種類還能看到紫外光。它們的聽覺同樣敏銳,這對於通過聲音交流和定位獵物至關重要。一些鳥類,如鴿子,甚至能夠感知地球的磁場,這有助於它們在長距離遷徙中進行導航。
鳥類的智力和學習能力也相當出眾。儘管它們的大腦相對較小,但結構緊湊,神經元密度高,使得某些鳥類能夠表現出複雜的認知能力,如工具使用、問題解決和社交學習。烏鴉科的鳥類尤其以其智慧聞名,它們能夠解決複雜的問題,甚至製作和使用簡單的工具。
繁殖方面,鳥類通過產卵繁衍後代。它們的育兒行為多種多樣,從完全獨立的早熟型幼鳥到需要長期照顧的晚熟型幼鳥都有。許多鳥類築巢、孵卵並餵養幼鳥,展現出複雜的親代照顧行為。
鳥類在生態系統中扮演著重要角色。作為捕食者,它們幫助控制昆蟲和小型動物的數量;作為種子傳播者,它們促進植物的繁衍和分佈;作為授粉者,某些鳥類如蜂鳥對植物的繁殖至關重要。此外,鳥類還是許多生態系統中能量和養分流動的重要環節。
鳥類的分佈幾乎遍及全球,從炎熱的熱帶雨林到寒冷的極地地區,從海洋到沙漠,都能看到鳥類的身影。它們通過各種適應性特徵來適應不同的環境,如企鵝的流線型身體適應水中生活,沙漠鳥類的特殊代謝機制幫助它們在乾旱環境中生存。
許多鳥類種類具有遷徙行為,每年在繁殖地和越冬地之間進行長距離飛行。這種行為不僅展示了鳥類驚人的耐力和導航能力,也反映了它們對季節性資源變化的適應。
總的來說,鳥類是一類獨特而多樣的動物,它們通過飛行能力、高效的生理系統、複雜的行為和廣泛的生態適應性,在地球上的幾乎每個角落都佔據了重要的生態位置。它們不僅在生態系統中發揮著關鍵作用,也因其美麗的外表、動聽的鳴叫和有趣的行為而深受人類喜愛,在人類文化中佔有重要地位。
1.2 鳥類的生態重要性
鳥類在生態系統中扮演著至關重要的角色,其生態重要性體現在多個方面。首先,作為食物網的重要組成部分,鳥類在維持生態平衡中發揮著關鍵作用。它們既是捕食者,也是被捕食者,參與調節各種生物的數量。許多鳥類以昆蟲為食,有效控制害蟲數量,為農業和林業提供了天然的生物防治手段。例如,山雀科鳥類在森林生態系統中捕食大量毛毛蟲,有助於保護樹木免受過度啃食。同時,猛禽如鷹和貓頭鷹則控制鼠類等小型哺乳動物的數量,間接影響整個生態系統的結構和功能。
鳥類在植物繁衍和分佈方面也起著重要作用。許多鳥類是重要的種子傳播者,通過消化系統或外部攜帶的方式將種子傳播到遠處。這種行為不僅有助於植物擴大分佈範圍,還促進了基因交流,增加了植物種群的遺傳多樣性。例如,歐洲松鴉在森林更新和擴張中發揮著重要作用,它們會將橡樹的果實埋藏在地下作為食物儲備,而那些未被取食的橡果最終可能發芽生長成新的樹木。在熱帶雨林中,大型鳥類如犀鳥可以將大顆種子傳播到遠處,對維持森林的多樣性和更新至關重要。
某些鳥類,特別是蜂鳥、太陽鳥和某些鸚鵡物種,是重要的授粉者。它們在採集花蜜的過程中,將花粉從一朵花傳播到另一朵花,促進了植物的有性繁殖。在一些生態系統中,鳥類甚至成為某些植物物種唯一的授粉媒介。例如,在南美洲的安第斯山脈,某些蜂鳥物種與特定的花卉物種形成了共生關係,這些花卉的形狀和顏色專門適應了蜂鳥的喙部結構和視覺特徵。
鳥類還通過其排泄物參與生態系統的養分循環。海鳥的糞便富含氮和磷,對於海島和海岸生態系統的養分輸入至關重要。在一些貧瘠的生態系統中,鳥類糞便可能是主要的養分來源,支持著豐富的植物和動物群落。例如,在秘魯的一些海岸地區,海鳥糞便(鳥糞石)的堆積形成了獨特的生態環境,支持著特有的植物和動物群落。
鳥類的遷徙行為對於不同生態系統之間的能量和物質流動具有重要意義。每年,數十億只鳥類在繁殖地和越冬地之間往返,在這個過程中,它們將養分和能量從一個生態系統轉移到另一個生態系統。例如,從北極繁殖地遷徙到南半球越冬的鷸鴴類鳥類,將大量的生物量和養分從北極地區帶到了熱帶和溫帶地區。
在水生生態系統中,鳥類也發揮著重要作用。涉禽如鷺鷥和鸛通過捕食魚類和兩棲動物,調節水生生物的數量。同時,它們在不同水體之間的移動也促進了水生生物的基因交流。水鳥如鴨子和天鵝通過攜帶水生植物的種子或孢子,促進了水生植物的傳播。
鳥類還是環境健康的重要指標。由於鳥類對環境變化敏感,它們的種群動態和行為變化可以反映生態系統的健康狀況。例如,猛禽種群的衰退可能指示環境污染問題,而遷徙鳥類到達時間的變化則可能反映全球氣候變化的影響。
在森林生態系統中,啄木鳥科鳥類扮演著「生態系統工程師」的角色。它們在樹幹上鑿洞築巢,不僅為自己提供棲息地,還為其他鳥類和小型哺乳動物創造了棲息場所。這些樹洞在森林生態系統中形成了重要的微生境,增加了生物多樣性。
鳥類還通過其行為影響生態系統的結構和功能。例如,某些地面覓食的鳥類通過刨土尋找食物,改變了土壤結構,促進了養分循環和種子萌發。在草原生態系統中,一些鳥類物種通過選擇性採食某些植物物種,間接影響了植被的組成和結構。
此外,鳥類在維持珊瑚礁生態系統健康方面也發揮著作用。一些海鳥物種以捕食珊瑚礁魚類的天敵為生,從而間接保護了珊瑚礁魚類種群。同時,它們的排泄物為珊瑚礁提供了重要的養分輸入。
鳥類的生態重要性還體現在它們與其他生物之間的共生關係。例如,某些鳥類物種與大型哺乳動物形成了互利共生關係,它們幫助哺乳動物清理皮膚上的寄生蟲,同時獲得食物。在非洲草原上,牛椋鳥經常可以看到停在斑馬或角馬背上,這種關係不僅有利於哺乳動物的健康,也為鳥類提供了食物來源。
人類為鳥類的保護付出了巨大努力,其中一個重要原因就是認識到了鳥類的生態重要性。通過保護鳥類及其棲息地,我們實際上是在保護整個生態系統的健康和功能。鳥類保護區的設立不僅保護了鳥類,還為其他野生動植物提供了庇護所。同時,鳥類觀察作為一項重要的生態旅遊活動,為當地社區帶來經濟效益,進一步促進了生態保護意識的提高。
1.3 鳥類在人類文化中的地位
鳥類自古以來就在人類文化中佔據著重要地位,它們的存在深深地影響了人類的藝術、宗教、文學、神話和日常生活。鳥類的多樣性、優雅的外形、飛行能力和悅耳的鳴叫,使它們成為人類想像力和創造力的重要源泉。
在許多古代文明中,鳥類常被賦予神聖的地位。古埃及人將鷹頭人身的荷魯斯視為太陽神和法老的化身,象徵著王權和神性。在古希臘神話中,雅典娜的聖鳥是貓頭鷹,代表智慧和洞察力。北歐神話中的奧丁常常與兩隻烏鴉為伴,這些烏鴉被認為是他的眼睛和耳朵,為他帶來世界各地的消息。
中國文化中,鳥類同樣具有重要意義。鳳凰被視為德行、仁義和祥瑞的象徵,常與帝王權威聯繫在一起。仙鶴則代表長壽和高尚,經常出現在傳統繪畫和詩詞中。在日本文化中,鶴也被視為吉祥和長壽的象徵,常見於藝術作品和折紙藝術中。
在文學領域,鳥類一直是作家和詩人靈感的重要來源。英國浪漫主義詩人濟慈的《夜鶯頌》讚美了夜鶯的歌聲,將其與人類的生命和藝術聯繫在一起。中國古代詩人如李白、杜甫等人的作品中也常見鳥類意象,如「鳥宿池中樹,僧敲月下門」,以鳥類的存在烘托出寧靜幽美的意境。
宗教文化中,鳥類也扮演著重要角色。在基督教傳統中,鴿子象徵和平與聖靈。諾亞方舟的故事中,鴿子帶回橄欖枝,預示洪水退去。在伊斯蘭教中,某些聖訓提到鳥類的重要性,如先知穆罕默德教導信徒善待動物,包括鳥類。佛教經典中也常見鳥類,如金翅鳥被視為護法神。
在民間傳說和迷信中,鳥類常被賦予預示吉凶的能力。例如,在西方文化中,烏鴉常被視為不祥之兆,而在某些原住民文化中,烏鴉卻被尊為智慧的化身。中國傳統文化中,喜鵲被認為能帶來好運和喜事,「喜鵲登枝」是常見的吉祥圖案。
鳥類在視覺藝術中的地位同樣舉足輕重。從古埃及的壁畫到文藝復興時期的油畫,再到現代藝術,鳥類一直是藝術家們鍾愛的主題。中國的花鳥畫是傳統繪畫的重要門類,畫家們通過細膩的筆觸捕捉鳥類的神態和動作,傳達對自然的理解和感悟。日本的浮世繪中也常見鳥類題材,如歌川廣重的《名所江戶百景》系列中就有多幅描繪鳥類的作品。
在音樂領域,鳥類的鳴叫聲啟發了眾多作曲家。法國作曲家梅西安以鳥類鳴叫為靈感創作了多部作品,如《鳥的系列音樂會》。貝多芬的《田園交響曲》中模仿了鳥叫聲,表達了對大自然的讚美。中國傳統樂曲中也有不少以鳥為主題的作品,如古琴曲《平沙落雁》,描繪了雁群飛行和降落的景象。
在日常生活和語言中,鳥類也留下了深刻的印記。許多慣用語和諺語都與鳥類有關,如「鳥語花香」形容春天的美好景象,「如鷹展翅」比喻高遠的志向。英語中也有類似的表達,如「as free as a bird」(像鳥兒一樣自由)。這些表達方式反映了人們對鳥類特性的觀察和理解。
鳥類在現代社會中continues享有特殊地位。觀鳥活動成為一種廣受歡迎的休閒方式,吸引了眾多愛好者。這不僅促進了人們對自然的瞭解和欣賞,也推動了生態保護意識的提高。鳥類攝影成為一門專門的藝術,攝影師們通過鏡頭捕捉鳥類的瞬間美態,展現自然的奇妙。
在商業和設計領域,鳥類形象被廣泛應用。許多公司和組織選擇鳥類作為標誌,如美國國家橄欖球聯盟的費城老鷹隊。鳥類的形象也常見於產品設計、時尚服飾和室內裝飾中,為現代生活增添自然元素。
教育方面,鳥類常被用作教導兒童認識自然和生態的工具。兒童文學中充滿了以鳥類為主角的故事,這些故事不僅娛樂了孩子們,也傳遞了關於友誼、勇氣和環境保護的重要訊息。
在某些文化中,鳥類還與特定的社會身份或地位相聯繫。例如,在中世紀歐洲,獵鷹被視為貴族的象徵,獵鷹運動是上層社會的重要活動。在某些原住民文化中,特定鳥類的羽毛被用於製作儀式用品或裝飾物,象徵著社會地位和精神力量。
鳥類在人類文化中的地位不僅體現了人類對自然美的欣賞,也反映了人類對自由、智慧和精神追求的嚮往。通過藝術、文學和日常生活中對鳥類的描繪和運用,人類表達了對生命、自然和宇宙的理解和感悟。鳥類的存在豐富了人類的文化生活,為人類提供了無盡的靈感和想像空間,同時也提醒著人類保護自然、珍惜生命的重要性。
1.4 鳥類學研究的歷史
鳥類學研究的歷史可以追溯到人類文明的早期,但作為一門科學學科,它的發展主要始於18世紀。在這之前,人類對鳥類的認識主要來源於實用需求、宗教信仰和藝術表現。古代文明如埃及、希臘和中國都有關於鳥類的記載,這些早期的觀察和描述為後來的鳥類學奠定了基礎。
亞裡士多德可以被視為鳥類學的先驅之一。在他的著作《動物志》中,記錄了約170種鳥類,並對它們的行為和形態進行了詳細描述。這些早期的觀察雖然混雜著一些錯誤和迷信,但仍然為後世的研究提供了寶貴的參考。
中世紀時期,歐洲的鳥類知識主要集中在修道院中。修士們編纂了許多動物誌,其中包含了對鳥類的描述。這些描述常常混合了事實和想像,反映了當時的宗教觀念和有限的科學認知。同時,在阿拉伯世界,學者們也在進行鳥類研究,其中最著名的是阿爾-賈希茲(Al-Jahiz)的《動物書》,其中包含了大量關於鳥類的觀察。
文藝復興時期標誌著鳥類學研究的新階段。隨著印刷技術的發展和探險活動的增加,更多關於鳥類的知識被記錄和傳播。1555年,法國博物學家皮埃爾·貝隆(Pierre Belon)出版了《鳥類的歷史》,這被認為是第一本現代意義上的鳥類學著作。貝隆的工作開創了比較解剖學在鳥類研究中的應用,為後來的分類學奠定了基礎。
17世紀,約翰·雷(John Ray)和法蘭西斯·威洛比(Francis Willughby)的合作標誌著鳥類分類學的開端。他們的著作《鳥類學》(Ornithologiae)首次嘗試基於解剖學特徵對鳥類進行系統分類。這一工作為後來林奈的分類系統提供了重要參考。
18世紀是鳥類學發展的關鍵時期。瑞典博物學家卡爾·林奈(Carl Linnaeus)在其著作《自然系統》中建立了現代生物分類學的基礎,為鳥類命名和分類提供了統一的方法。林奈的二名法命名系統至今仍在使用,極大地推動了鳥類學的發展。
同時期,法國博物學家布封(Buffon)編纂的《自然史》中包含了大量關於鳥類的內容,其生動的描述和精美的插圖使鳥類學知識得到了廣泛傳播。布封的工作強調了環境對物種形成的影響,為後來的進化理論提供了啟發。
19世紀,鳥類學研究進入了快速發展期。查爾斯·達爾文(Charles Darwin)的進化論對鳥類學產生了深遠影響。達爾文在加拉帕戈斯群島觀察到的鳥類變異,尤其是達爾文雀的喙部差異,為他的自然選擇理論提供了重要證據。這一發現開啟了研究鳥類適應性進化的新篇章。
19世紀後半葉,鳥類學開始分化為多個專門領域。德國動物學家恩斯特·黑克爾(Ernst Haeckel)提出的生物發生律推動了鳥類胚胎學的發展。同時,鳥類行為學也開始興起,奧斯卡·海因羅特(Oskar Heinroth)和康拉德·洛倫茲(Konrad Lorenz)的開創性工作為這一領域奠定了基礎。
20世紀初,鳥類學研究方法不斷創新。鳥類環志技術的發明使得研究鳥類遷徙成為可能。埃爾溫·施特雷澤曼(Erwin Stresemann)的工作推動了鳥類系統分類學的發展,他強調了將解剖學、生態學和行為學等多學科方法結合應用於鳥類分類。
隨著技術的進步,鳥類學研究工具也不斷更新。望遠鏡和攝影技術的改進使得野外觀察變得更加精確。錄音設備的發展則為鳥類聲音研究開闢了新天地。這些技術進步大大擴展了鳥類學研究的範圍和深度。
二戰後,生態學在鳥類研究中的應用日益廣泛。大衛·拉克(David Lack)的開創性工作將生態學理論應用於鳥類種群研究,極大地推動了鳥類生態學的發展。同時,行為生態學的興起為理解鳥類行為提供了新的視角,尼科·廷伯根(Niko Tinbergen)等人的研究對這一領域貢獻巨大。
20世紀後半葉,分子生物學技術在鳥類學中的應用開闢了新的研究領域。DNA分析技術的應用使得研究鳥類的進化關係和遺傳多樣性成為可能。這些方法不僅revolutionized了鳥類分類學,也為研究鳥類的適應性進化和種群遺傳學提供了powerful工具。
近年來,鳥類保護生物學成為鳥類學的重要分支。隨著人類活動對自然環境影響的加劇,許多鳥類物種面臨滅絕威脅。鳥類學家們開始將研究重點轉向瀕危物種的保護,開發了一系列保護策略和管理方法。
同時,公民科學在鳥類學研究中的作用日益突出。大規模的鳥類普查項目,如聖誕節鳥類統計(Christmas Bird Count),為研究鳥類種群動態提供了寶貴數據。這種參與式研究不僅豐富了科學數據,也提高了公眾的環境意識。
當代鳥類學研究正朝著多學科交叉的方向發展。神經生物學、認知科學等領域的方法被引入鳥類研究,揭示了鳥類智力和學習能力的驚人之處。同時,全球氣候變化對鳥類的影響也成為研究熱點,鳥類被視為氣候變化的重要指示生物。
鳥類學的發展歷程反映了人類對自然認知的不斷深入。從早期的觀察描述,到現代的多學科綜合研究,鳥類學不斷拓展著我們對這些神奇生物的理解。今天,鳥類學不僅是一門純粹的自然科學,也與環境保護、生態平衡等全球性議題密切相關,在人類理解和保護地球生態系統的過程中發揮著重要作用。
第二章 鳥類多樣性與分類
2.1 鳥類的起源
鳥類是一個令人著迷的動物類群,其分類、命名和起源都有著豐富而複雜的歷史。
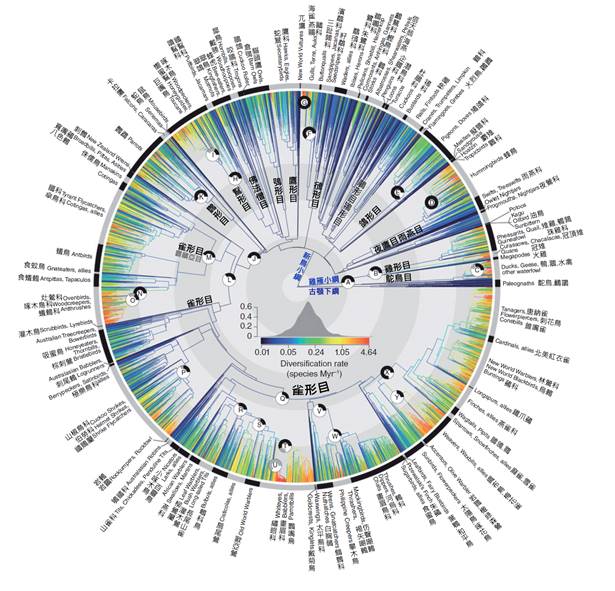
在現代生物分類系統中,鳥類被歸類為脊索動物門(Chordata)中的鳥綱(Aves)。這個分類反映了鳥類與其他脊椎動物的親緣關係,同時也突出了它們獨特的特徵。鳥綱下面還可以進一步細分為亞綱、目、科、屬和種等分類階層。目前,科學界認識到的現存鳥類約有10,000多種,分屬於約40個目。
其中最大的目是雀形目(Passeriformes),包括了鳥類總數的一半以上,如麻雀、燕子和烏鴉等。其他重要的目包括鸚鵡目(Psittaciformes)、鴿形目(Columbiformes)和鶴形目(Gruiformes)等。每個目都有其獨特的特徵和生態位,反映了鳥類在進化過程中的多樣化適應。
鳥類的命名遵循國際動物命名法規(ICZN),採用二名法。每個鳥類物種都有一個由屬名和種加詞組成的學名,例如家鴿的學名是(Columba livia)。這種命名方式不僅提供了一個統一的國際交流平臺,也反映了物種間的親緣關係。
除了學名,鳥類還有各種俗名或通用名。這些名稱通常基於鳥類的外觀、行為或棲息地等特徵,在不同的文化和語言中可能有很大差異。例如,同一種鳥在中文中可能叫"喜鵲",而在英語中則稱為"Magpie"。
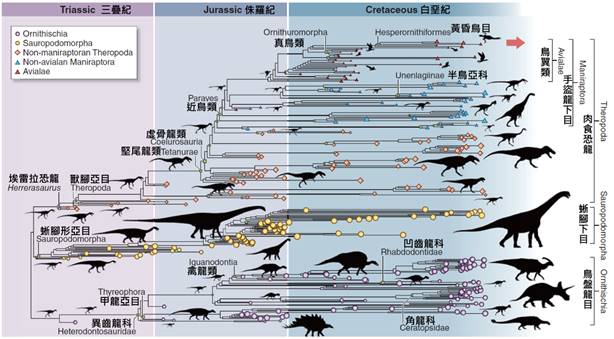
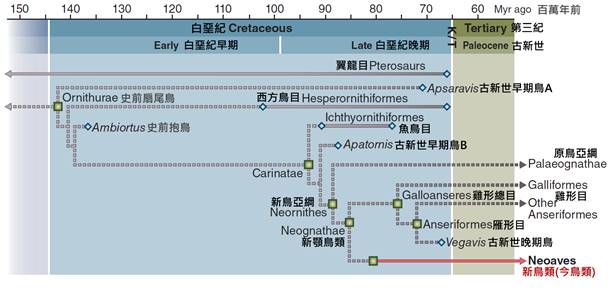
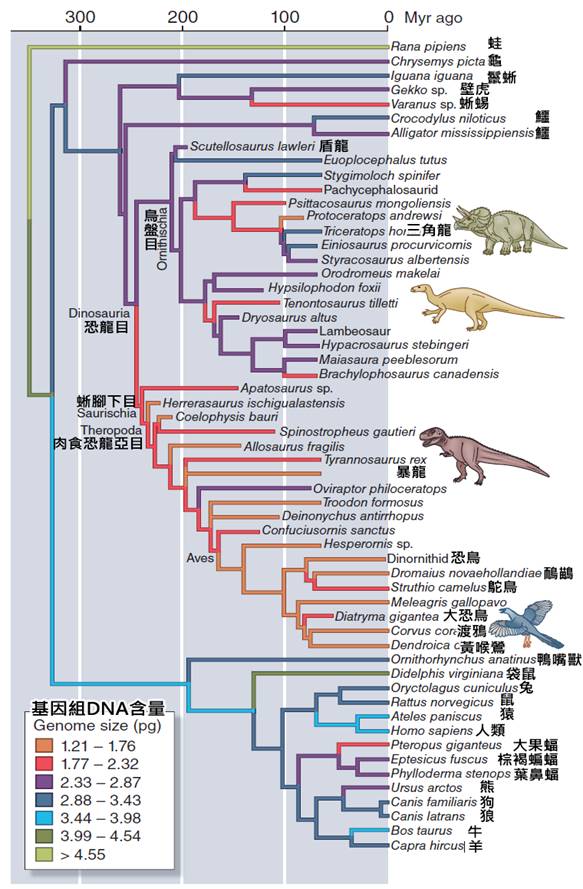
關於鳥類的起源,這是一個長期以來吸引科學家關注的話題。現代古生物學和分子生物學的研究表明,鳥類起源於中生代晚侏羅紀時期,大約1.5億至2億年前。它們是從一群被稱為獸腳類的恐龍演化而來的。
最著名的過渡化石是始祖鳥(Archaeopteryx),發現於德國的侏羅紀地層中。始祖鳥既有爬行動物的特徵,如長尾和有爪的前肢,又具有鳥類的特徵,如羽毛和叉骨。這個化石為鳥類起源於恐龍提供了重要證據。
隨著更多化石的發現和研究,科學家們逐漸勾勒出了鳥類演化的大致輪廓。在晚侏羅紀和早白堊紀期間,一些小型肉食恐龍逐漸演化出了一些鳥類的特徵,如羽毛、中空骨骼和叉骨等。這些特徵最初可能是為了保暖或求偶展示,後來才逐漸適應了飛行。
飛行能力的獲得是鳥類演化史上的一個重要里程碑。它不僅改變了鳥類的生態位,也推動了它們的多樣化。隨著飛行能力的完善,鳥類逐漸colonized了各種生態環境,從熱帶雨林到極地冰原,從海洋到沙漠。
值得注意的是,鳥類的演化並非一蹴而就,而是一個漫長而複雜的過程。在這個過程中,許多原始鳥類支系出現又滅絕,最終形成了我們今天所見的現代鳥類。例如,在白堊紀末期的大滅絕事件中,許多恐龍和原始鳥類滅絕了,但一些鳥類倖存下來,並在之後的地質時期中迅速輻射演化,形成了現代鳥類的主要類群。
現代分子生物學和基因組學研究為鳥類的演化提供了新的視角。通過比較不同鳥類物種的DNA序列,科學家們可以重建鳥類的演化樹,並估算不同類群分化的時間。這些研究不僅confirmed了化石記錄的發現,還揭示了許多化石難以展示的演化細節。
鳥類的分類、命名和起源研究不僅對於理解生物多樣性有重要意義,也為我們理解生命演化的general規律提供了寶貴的案例。隨著新技術和新方法的不斷發展,我們對鳥類的認識也在不斷深化。例如,近年來的研究顯示,鳥類可能比我們之前認為的更聰明,許多種類具有使用工具、解決問題甚至自我認知的能力。
鳥類的研究也為保護生物多樣性提供了重要依據。通過瞭解不同鳥類物種的分類地位和演化歷史,我們可以更好地制定保護策略,特別是對於那些瀕危物種。同時,鳥類作為許多生態系統的重要成員,其分佈和數量的變化often可以反映環境變化,因此在生態監測中扮演著重要角色。
儘管我們對鳥類的認識已經相當豐富,但仍有許多問題待解。例如,一些鳥類類群的演化關係仍存在爭議,一些古老支系的生態位和生活方式也還不清楚。此外,隨著氣候變化和人類活動的影響,鳥類面臨著新的演化壓力,它們如何應對這些變化也是一個值得關注的問題。
2.2 鳥類的演化樹
鳥類的分類、命名和物種親緣學是生物學研究中一個引人入勝的領域,它不僅揭示了鳥類的多樣性,還闡明瞭它們之間的演化關係。讓我們從鳥類的分類開始,逐步深入探討這個主題。
在現代生物分類系統中,鳥類被歸類為脊索動物門(Chordata)下的鳥綱(Aves)。這個分類反映了鳥類與其他脊椎動物的關係,同時也突出了它們的獨特特徵。鳥綱下可以進一步細分為亞綱、目、科、屬和種等分類階層。目前,科學界認識到的現存鳥類約有10,000多種,分屬於約40個目。
其中最大的目是雀形目(Passeriformes),包括了鳥類總數的一半以上,如麻雀、燕子和烏鴉等。其他重要的目包括鸚鵡目(Psittaciformes)、鴿形目(Columbiformes)和鶴形目(Gruiformes)等。每個目都有其獨特的特徵和生態位,反映了鳥類在演化過程中的多樣化適應。
鳥類的命名遵循國際動物命名法規(ICZN),採用二名法。每個鳥類物種都有一個由屬名和種加詞組成的學名,例如家鴿的學名是(Columba livia)。這種命名方式不僅提供了一個統一的國際交流平臺,也反映了物種間的親緣關係。除了學名,鳥類還有各種俗名或通用名,這些名稱通常基於鳥類的外觀、行為或棲息地等特徵,在不同的文化和語言中可能有很大差異。
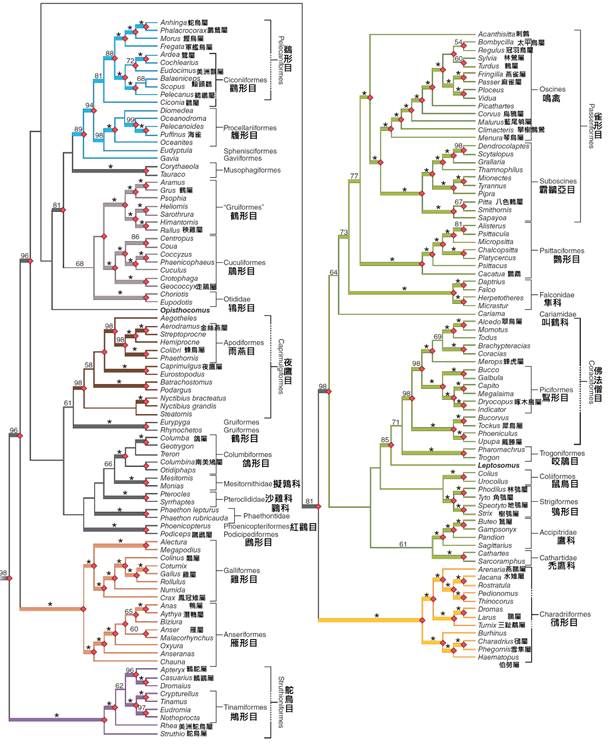
物種的親緣學是研究生物之間演化關係的學科,而鳥類的演化樹則是描述鳥類親緣關係的一種圖形表示。這棵「樹」的每個分支代表一個演化譜系,分支的分叉點代表物種分化事件。通過構建演化樹,科學家們可以更好地理解鳥類的演化歷史和物種間的關係。
鳥類的演化樹的構建是一個複雜而持續的過程,涉及多個科學領域的交叉。傳統上,科學家們主要依賴形態學特徵來建立親緣關係。他們通過比較不同鳥類物種的外部形態(如喙的形狀、羽毛的結構)和內部解剖結構(如骨骼、肌肉系統)來推斷它們之間的關係。
然而,隨著分子生物學技術的發展,DNA序列比較成為構建演化樹的重要工具。通過比較不同鳥類物種的基因或蛋白質序列,科學家們可以更精確地估算物種間的親緣關係和分化時間。這種方法的優勢在於它可以揭示形態學方法難以檢測的微小差異,並且能夠提供關於演化速率的資訊。
近年來,隨著高通量測序技術的進步,全基因組比較分析成為研究鳥類親緣關係的新趨勢。這種方法可以同時分析數千個基因,提供更全面、更準確的親緣關係估計。例如,2014年發表的鳥類基因組計劃(Avian Phylogenomics Project)分析了48個鳥類物種的全基因組,大大改進了我們對鳥類高級分類群之間關係的理解。
鳥類的演化樹研究不僅揭示了現存鳥類之間的關係,還幫助我們理解了鳥類的起源和早期演化。現代研究表明,鳥類起源於中生代晚侏羅紀時期,約1.5億至2億年前,它們是從一群被稱為獸腳類的恐龍演化而來的。最著名的過渡化石是始祖鳥(Archaeopteryx),它既有爬行動物的特徵,又具有鳥類的特徵,為鳥類起源於恐龍提供了重要證據。
鳥類的演化樹顯示,現代鳥類可以分為兩大類群:古頜總目(Palaeognathae)和今頜總目(Neognathae)。古頜總目包括鴕鳥、鵜鶘等不會飛的鳥類,而今頜總目則包括了絕大多數的現代鳥類。
在今頜總目中,水禽類(Galloanserae)是一個重要的分支,包括鴨、鵝、雞等。另一個主要分支是新新鳥類(Neoaves),包括了從猛禽到鸚鵡、從海鳥到雀形目等多樣化的鳥類。雀形目是新新鳥類中最大的一個目,也是整個鳥類中最為多樣化的一組,包括了約6,000種鳥類。
鳥類演化樹的研究還揭示了一些令人驚訝的親緣關係。例如,鸚鵡和鷹隼類被發現是近親,儘管它們在形態和生活習性上差異很大。這種「意外」的親緣關係提醒我們,演化過程中的趨同演化和趨異演化可能會導致形態特徵與真實親緣關係的不一致。
鳥類的演化樹研究還幫助我們理解了鳥類的適應性輻射。例如,達爾文的雀(Darwin's finches)在加拉帕戈斯群島上的輻射演化是一個經典案例,展示了how鳥類如何在相對短的時間內適應不同的生態位。通過研究這些鳥類的演化樹,科學家們可以追蹤特定形態特徵(如喙的形狀)的演化歷史。
此外,鳥類演化樹的研究還為我們理解生物地理學提供了重要線索。通過比較不同地理區域鳥類的親緣關係,科學家們可以推斷古代大陸的分離和連接歷史,以及鳥類的遷徙和擴散模式。
然而,儘管取得了巨大進展,鳥類的演化樹仍然存在一些爭議和未解之謎。例如,新新鳥類的早期分化過程仍然不是很清楚,這可能是由於這些鳥類在很短的時間內迅速分化,留下的遺傳信號較弱。此外,一些特殊類群(如被稱為「生活化石」的冠鳩)的演化位置也仍有爭議。
鳥類的分類、命名和演化樹研究不僅對於理解生物多樣性有重要意義,也為我們理解生命演化的一般規律提供了寶貴的案例。這些研究結果對於生物多樣性保護也有重要應用。通過瞭解物種間的親緣關係,我們可以更好地評估生物多樣性的價值,並制定更有效的保護策略。
隨著新技術和新方法的不斷發展,我們對鳥類的認識也在不斷深化。例如,單細胞測序技術可能會讓我們更深入地瞭解鳥類基因組的演化過程。同時,古DNA技術的進步可能會讓我們有機會研究已滅絕鳥類的基因組,從而填補演化樹上的一些空白。
鳥類的演化樹研究還為我們提供了一個理解大尺度演化過程的窗口。通過研究鳥類如何在不同環境中適應和分化,我們可以獲得關於物種形成、適應性演化和生態位分化的深刻見解。這些知識不僅對於理解過去的演化歷史很重要,也有助於我們預測鳥類如何應對未來的環境變化。
2.3 鳥類的演化歷史
鳥類的演化歷史是一個複雜而引人入勝的過程,這個過程揭示了生命如何從地面爬行動物演變為天空的主宰者。鳥類的起源可以追溯到中生代三疊紀末期,大約兩億兩千萬年前。在這個時期,一群被稱為獸腳類恐龍的小型肉食恐龍開始了向鳥類演化的旅程。
這些早期的獸腳類恐龍具有一些鳥類的原始特徵,如中空的骨骼和三趾腳。隨著時間的推移,這些特徵逐漸演變,使得這些生物更加適應於攀爬和滑翔。在侏羅紀早期,約兩億年前,出現了一些更接近鳥類的恐龍,如始祖鳥。這些生物已經具備了一些鳥類的關鍵特徵,如羽毛和翼狀前肢。
羽毛的演化是鳥類發展史上的一個重要里程碑。最早的羽毛可能只是簡單的絲狀結構,主要用於保溫。隨著時間的推移,羽毛變得更加複雜,開始具備空氣動力學功能。這一過程為後來的飛行能力奠定了基礎。值得注意的是,許多非鳥類恐龍也有羽毛,這表明羽毛的出現早於真正的鳥類。
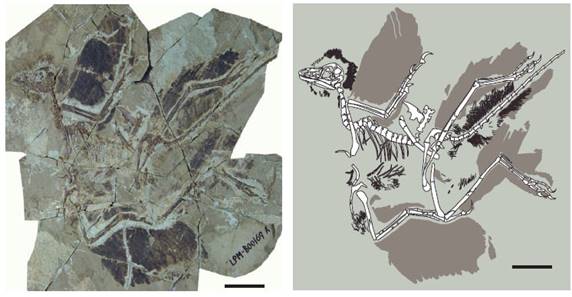
在侏羅紀晚期,約一億五千萬年前,出現了被認為是現代鳥類直接祖先的始祖鳥。始祖鳥是一個關鍵的過渡物種,它既有爬行動物的特徵(如長尾和有齒的喙),也有鳥類的特徵(如羽毛和翼)。始祖鳥的發現填補了爬行動物和鳥類之間的演化空白,為鳥類起源於恐龍提供了有力證據。
在白堊紀時期,約一億四千萬年前到六千五百萬年前,鳥類經歷了快速的多樣化。這個時期出現了許多早期鳥類群,如反鳥類和真鳥類。反鳥類保留了一些原始特徵,如有齒的喙和爪狀的翼指,而真鳥類則更接近現代鳥類。這個時期的化石記錄顯示,鳥類開始適應各種生態位,包括水生環境和陸地環境。
飛行能力的演化是鳥類成功的關鍵。最初的飛行可能是從高處滑翔開始的,隨後發展出拍打翅膀的能力。飛行肌肉的發達、骨骼的輕質化、以及呼吸系統的改進都是為了支持飛行而發生的重要演化。這些適應使鳥類能夠有效地利用空中生態位,這在當時是一個相對未被占據的領域。
白堊紀末期的大滅絕事件(約六千五百萬年前)對鳥類的演化產生了深遠影響。這次滅絕事件導致了包括非鳥類恐龍在內的許多物種的消失,但一些鳥類成功地度過了這個危機。這些倖存的鳥類成為了現代鳥類的祖先。滅絕事件後的生態空缺為鳥類的快速輻射適應創造了條件。
在新生代,鳥類經歷了爆炸式的演化。早期的現代鳥類開始分化為我們今天看到的主要類群。水禽類、陸禽類、鴿形目、鸚鵡目等多種類群在這個時期出現並快速分化。這種多樣化反映了鳥類適應不同環境和生態位的能力。
鳥類的大腦也在演化過程中發生了顯著變化。相比其他脊椎動物,鳥類的大腦相對體型更大,特別是負責高級認知功能的區域。這種演化使得許多鳥類具備了複雜的社會行為、工具使用能力和高度發達的學習能力。
鳥類的發聲器官 - 鳴管 - 的演化是另一個重要的適應。鳴管的發展使得鳥類能夠產生複雜的聲音,這對於交流、領地宣示和求偶至關重要。不同鳥類種類的鳴叫能力差異很大,反映了它們在聲音通訊方面的不同演化策略。
遷徙能力的演化是鳥類適應季節性資源變化的重要策略。許多鳥類發展出長距離遷徙的能力,這需要複雜的導航系統、高效的能量利用和適應不同環境的能力。遷徙行為的演化極大地擴展了鳥類的地理分佈範圍。
島嶼環境在鳥類演化中扮演了特殊角色。由於島嶼的隔離性,許多獨特的鳥類種類在這些環境中演化而來。達爾文在加拉帕戈斯群島觀察到的地雀就是一個著名的例子,展示了鳥類如何通過適應性輻射來適應不同的生態位。
鳥類的體型在演化過程中也發生了顯著變化。從早期的較大型鳥類到現代的各種體型,鳥類展現了極大的多樣性。最小的蜂鳥到最大的鴕鳥,體現了鳥類對不同生態位的適應。
值得注意的是,一些鳥類類群在演化過程中失去了飛行能力。例如,鴕鳥、企鵝等鳥類為了適應特定環境,犧牲了飛行能力而發展出其他特化的能力。這種演化趨勢反映了自然選擇如何塑造物種以適應特定的生態位。
現代分子生物學技術為研究鳥類演化提供了新的工具。DNA分析揭示了許多之前未知的演化關係,有時甚至挑戰了基於形態學的傳統分類。例如,基因研究顯示鸚鵡與鳩鴿類的親緣關係比之前認為的要近。
氣候變化一直是驅動鳥類演化的重要因素。過去的氣候波動導致了棲息地的變化,促使鳥類適應新的環境。例如,冰河時期的氣候變化影響了許多鳥類的分佈和演化軌跡。
人類活動對鳥類的演化產生了深遠影響。棲息地破壞、過度捕獵和引入外來物種等因素導致了許多鳥類物種的滅絕,同時也為一些適應性強的物種創造了新的機會。這種人為選擇壓力正在加速某些鳥類的演化速度。
鳥類的演化歷史是一個持續的過程。即使在當代,我們仍然可以觀察到鳥類在適應新環境和應對新挑戰方面的演化。例如,一些城市鳥類正在發展出應對噪音污染的新行為和生理適應。這些當代的演化例子提醒我們,演化是一個持續的、動態的過程。
從恐龍到鳥類的演化過程是古生物學和進化生物學中最引人入勝的故事之一。這個轉變過程橫跨數千萬年,涉及一系列複雜的形態學和生理學變化,最終導致了現代鳥類的出現。這個過程不僅揭示了生命的適應性和韌性,也為我們理解進化機制提供了寶貴的洞察。
這個故事始於晚三疊紀,大約2.3億年前。當時,一群被稱為獸腳類的恐龍開始了向鳥類演化的漫長旅程。這些早期的獸腳類恐龍,如鐮刀龍,已經具備了一些鳥類的原始特徵,比如中空的骨骼和三趾腳。這些特徵為後來的飛行適應奠定了基礎。
隨著時間的推移,獸腳類恐龍逐漸演化出更多鳥類特徵。在侏羅紀早期,約2億年前,出現了更接近鳥類的恐龍類群,如曼努恐龍科。這些恐龍開始發展出羽毛樣結構,這是向鳥類過渡的關鍵步驟。最初,這些原始羽毛可能主要用於保溫或求偶展示,而非飛行。
羽毛的演化是從恐龍到鳥類轉變過程中的一個重要里程碑。最早的羽毛可能只是簡單的絲狀結構,類似於現代鳥類的絨羽。隨著時間的推移,羽毛變得更加複雜,逐漸發展出具有中軸和羽枝的結構。這種結構為後來的飛行羽毛奠定了基礎。值得注意的是,許多非鳥類恐龍也有羽毛,這表明羽毛的出現早於真正的飛行能力。
在侏羅紀晚期,約1.5億年前,出現了被認為是現代鳥類直接祖先的始祖鳥。始祖鳥是一個關鍵的過渡物種,它同時具備了爬行動物和鳥類的特徵。它保留了恐龍的一些特徵,如長骨骼尾和有齒的喙,但同時也發展出了鳥類的特徵,如羽毛覆蓋的身體和翼狀前肢。始祖鳥的發現為鳥類起源於恐龍提供了決定性的證據。
從始祖鳥到現代鳥類的演化過程涉及一系列重要的解剖學變化。其中最顯著的是骨骼系統的改變。恐龍的長骨骼尾逐漸縮短,形成了鳥類特有的尾綱骨。前肢逐漸演變成適合飛行的翅膀,手指融合,形成了翼尖。後肢也發生了變化,適應了抓握和站立的需求。
頭骨的演化也是一個重要方面。恐龍的堅硬下頜逐漸演變成鳥類的輕巧喙部。牙齒逐漸消失,被角質喙所取代。這種變化不僅減輕了頭部的重量,也提高了取食效率。同時,頭骨變得更加輕盈,眼眶擴大,這些都有利於飛行生活。
內部器官系統也經歷了重大變革。最顯著的是呼吸系統的演化。恐龍的簡單肺部逐漸發展成鳥類特有的高效氣囊系統。這種系統允許空氣在肺部單向流動,大大提高了氧氣吸收效率,為高強度飛行活動提供了必要的能量支援。
心臟血管系統也發生了重要變化。鳥類發展出了四腔心臟,能夠完全分離含氧和缺氧血液。這種適應提高了新陳代謝效率,為飛行提供了持續的能量供應。同時,鳥類的體溫調節系統也變得更加高效,發展出了恆溫特性。
神經系統,特別是大腦的演化,是從恐龍到鳥類轉變過程中另一個引人注目的方面。鳥類的大腦相對體型更大,特別是負責高級認知功能的區域。這種演化使得鳥類具備了複雜的學習能力、社會行為和問題解決能力。
生殖系統也經歷了重要變化。恐龍的雙側輸卵管逐漸演化為鳥類的單側輸卵管。這種變化可能與減輕飛行時的體重有關。同時,蛋殼的結構也發生了變化,變得更加堅固yet透氣,更適合在地面環境中孵化。
行為和生活方式的演化也是這個過程的重要組成部分。早期的飛行可能是從高處滑翔開始的,隨後才發展出主動飛行能力。這種行為上的改變推動了形態學適應的進一步發展,如飛行肌肉的增強和骨骼的進一步輕質化。
值得注意的是,從恐龍到鳥類的演化並非一個線性過程。在這個漫長的時期內,出現了許多具有鳥類特徵的恐龍類群,如德荷鳥、小盜龍等。這些類群展示了不同程度的"鳥化"特徵,反映了演化過程的複雜性和多樣性。
白堊紀末期的大滅絕事件(約6600萬年前)對這個演化過程產生了深遠影響。這次事件導致了包括非鳥類恐龍在內的許多物種的滅絕,但一些早期鳥類成功地度過了這個危機。這些倖存的鳥類成為了現代鳥類的祖先。滅絕事件後的生態空缺為鳥類的快速輻射適應創造了條件。
在新生代,鳥類經歷了爆炸式的演化和多樣化。早期的現代鳥類開始分化為我們今天看到的主要類群。這種多樣化反映了鳥類適應各種生態位的能力,從海洋到沙漠,從熱帶雨林到極地苔原,鳥類成功地占據了幾乎所有的陸地和水生環境。
近年來,分子生物學和基因組學研究為理解從恐龍到鳥類的演化提供了新的視角。這些研究不僅證實了鳥類與獸腳類恐龍的密切關係,也揭示了許多形態學和生理學變化背後的遺傳基礎。例如,研究發現控制喙部形態的基因在非鳥類獸腳類恐龍中就已存在,這解釋了為什麼某些恐龍具有類似鳥喙的結構。
從恐龍到鳥類的演化過程是生命適應性和韌性的絕佳例證。它展示了自然選擇如何塑造生物以適應新的生態位,同時也揭示了生命形式的驚人連續性。這個過程不僅改變了我們對恐龍和鳥類的理解,也深化了我們對進化機制的認識,為現代生物學研究提供了豐富的洞察和啟示。
2.4 從恐龍到鳥類
這個故事始於晚三疊紀,大約2.3億年前。當時,一群被稱為獸腳類的恐龍開始了向鳥類演化的漫長旅程。這些早期的獸腳類恐龍,如鐮刀龍,已經具備了一些鳥類的原始特徵,比如中空的骨骼和三趾腳。這些特徵為後來的飛行適應奠定了基礎。
隨著時間的推移,獸腳類恐龍逐漸演化出更多鳥類特徵。在侏羅紀早期,約2億年前,出現了更接近鳥類的恐龍類群,如曼努恐龍科。這些恐龍開始發展出羽毛樣結構,這是向鳥類過渡的關鍵步驟。最初,這些原始羽毛可能主要用於保溫或求偶展示,而非飛行。
羽毛的演化是從恐龍到鳥類轉變過程中的一個重要里程碑。最早的羽毛可能只是簡單的絲狀結構,類似於現代鳥類的絨羽。隨著時間的推移,羽毛變得更加複雜,逐漸發展出具有中軸和羽枝的結構。這種結構為後來的飛行羽毛奠定了基礎。值得注意的是,許多非鳥類恐龍也有羽毛,這表明羽毛的出現早於真正的飛行能力。
在侏羅紀晚期,約1.5億年前,出現了被認為是現代鳥類直接祖先的始祖鳥。始祖鳥是一個關鍵的過渡物種,它同時具備了爬行動物和鳥類的特徵。它保留了恐龍的一些特徵,如長骨骼尾和有齒的喙,但同時也發展出了鳥類的特徵,如羽毛覆蓋的身體和翼狀前肢。始祖鳥的發現為鳥類起源於恐龍提供了決定性的證據。
從始祖鳥到現代鳥類的演化過程涉及一系列重要的解剖學變化。其中最顯著的是骨骼系統的改變。恐龍的長骨骼尾逐漸縮短,形成了鳥類特有的尾綱骨。前肢逐漸演變成適合飛行的翅膀,手指融合,形成了翼尖。後肢也發生了變化,適應了抓握和站立的需求。
頭骨的演化也是一個重要方面。恐龍的堅硬下頜逐漸演變成鳥類的輕巧喙部。牙齒逐漸消失,被角質喙所取代。這種變化不僅減輕了頭部的重量,也提高了取食效率。同時,頭骨變得更加輕盈,眼眶擴大,這些都有利於飛行生活。
內部器官系統也經歷了重大變革。最顯著的是呼吸系統的演化。恐龍的簡單肺部逐漸發展成鳥類特有的高效氣囊系統。這種系統允許空氣在肺部單向流動,大大提高了氧氣吸收效率,為高強度飛行活動提供了必要的能量支援。
心臟血管系統也發生了重要變化。鳥類發展出了四腔心臟,能夠完全分離含氧和缺氧血液。這種適應提高了新陳代謝效率,為飛行提供了持續的能量供應。同時,鳥類的體溫調節系統也變得更加高效,發展出了恆溫特性。
神經系統,特別是大腦的演化,是從恐龍到鳥類轉變過程中另一個引人注目的方面。鳥類的大腦相對體型更大,特別是負責高級認知功能的區域。這種演化使得鳥類具備了複雜的學習能力、社會行為和問題解決能力。
生殖系統也經歷了重要變化。恐龍的雙側輸卵管逐漸演化為鳥類的單側輸卵管。這種變化可能與減輕飛行時的體重有關。同時,蛋殼的結構也發生了變化,變得更加堅固yet透氣,更適合在地面環境中孵化。
行為和生活方式的演化也是這個過程的重要組成部分。早期的飛行可能是從高處滑翔開始的,隨後才發展出主動飛行能力。這種行為上的改變推動了形態學適應的進一步發展,如飛行肌肉的增強和骨骼的進一步輕質化。
值得注意的是,從恐龍到鳥類的演化並非一個線性過程。在這個漫長的時期內,出現了許多具有鳥類特徵的恐龍類群,如德荷鳥、小盜龍等。這些類群展示了不同程度的"鳥化"特徵,反映了演化過程的複雜性和多樣性。
白堊紀末期的大滅絕事件(約6600萬年前)對這個演化過程產生了深遠影響。這次事件導致了包括非鳥類恐龍在內的許多物種的滅絕,但一些早期鳥類成功地度過了這個危機。這些倖存的鳥類成為了現代鳥類的祖先。滅絕事件後的生態空缺為鳥類的快速輻射適應創造了條件。
在新生代,鳥類經歷了爆炸式的演化和多樣化。早期的現代鳥類開始分化為我們今天看到的主要類群。這種多樣化反映了鳥類適應各種生態位的能力,從海洋到沙漠,從熱帶雨林到極地苔原,鳥類成功地占據了幾乎所有的陸地和水生環境。
近年來,分子生物學和基因組學研究為理解從恐龍到鳥類的演化提供了新的視角。這些研究不僅證實了鳥類與獸腳類恐龍的密切關係,也揭示了許多形態學和生理學變化背後的遺傳基礎。例如,研究發現控制喙部形態的基因在非鳥類獸腳類恐龍中就已存在,這解釋了為什麼某些恐龍具有類似鳥喙的結構。
從恐龍到鳥類的演化過程是生命適應性和韌性的絕佳例證。它展示了自然選擇如何塑造生物以適應新的生態位,同時也揭示了生命形式的驚人連續性。這個過程不僅改變了我們對恐龍和鳥類的理解,也深化了我們對進化機制的認識,為現代生物學研究提供了豐富的洞察和啟示。



2.5 鳥類的命名
鳥類是一個多樣性極其豐富的動物類群,其分類和命名體系反映了這種多樣性,同時也為我們理解鳥類的演化歷史和生態適應提供了重要框架。讓我們深入探討鳥類的分類、命名以及多樣性分類的各個方面。
在現代生物分類系統中,鳥類被歸類為脊索動物門(Chordata)中的鳥綱(Aves)。這個分類反映了鳥類與其他脊椎動物的親緣關係,同時也突出了它們獨特的特徵。鳥綱下面還可以進一步細分為亞綱、目、科、屬和種等分類階層。目前,科學界認識到的現存鳥類約有10,000多種,分屬於約40個目。
鳥類的命名遵循國際動物命名法規(ICZN),採用二名法。每個鳥類物種都有一個由屬名和種加詞組成的學名,例如家鴿的學名是(Columba livia)。這種命名方式不僅提供了一個統一的國際交流平臺,也反映了物種間的親緣關係。除了學名,鳥類還有各種俗名或通用名。這些名稱通常基於鳥類的外觀、行為或棲息地等特徵,在不同的文化和語言中可能有很大差異。
鳥類多樣性的分類是一個複雜而持續演進的過程。傳統上,鳥類學家主要根據形態特徵來劃分鳥類類群。然而,隨著分子生物學技術的發展,DNA序列分析在鳥類分類中發揮了越來越重要的作用。這種方法能夠揭示形態學難以檢測的隱蔽種,並且更準確地反映物種間的演化關係。
在現代鳥類分類系統中,鳥類首先被分為兩個主要類群:古頜總目(Palaeognathae)和今頜總目(Neognathae)。古頜總目包括鴕鳥、鵜鶘等不會飛的鳥類,而今頜總目則包括了絕大多數的現代鳥類。
古頜總目是一個相對較小的類群,包括了鴕鳥目(Struthioniformes)、鵜鶘目(Tinamiformes)等。這些鳥類通常體型較大,不能飛行,主要分佈在南半球。它們保留了許多原始特徵,被認為是現代鳥類中最早分化出來的群體。
今頜總目則包含了絕大多數的現代鳥類。在這個類群中,水禽類(Galloanserae)是一個重要的分支,包括鴨、鵝、雞等。另一個主要分支是新新鳥類(Neoaves),包括了從猛禽到鸚鵡、從海鳥到雀形目等多樣化的鳥類。
雀形目(Passeriformes)是新新鳥類中最大的一個目,也是整個鳥類中最為多樣化的一組,包括了約6,000種鳥類,佔所有鳥類物種的一半以上。這個目包括了我們常見的許多鳥類,如麻雀、燕子、烏鴉等。雀形目鳥類的多樣性體現在它們的形態、行為和生態位上,從體型微小的蜂鳥到體型較大的烏鴉,從專門捕食昆蟲的鶯科鳥類到主要食用種子的雀科鳥類,都屬於這個目。
鸚鵡目(Psittaciformes)是另一個引人注目的鳥類類群。這個目包括了鸚鵡、鸚哥等鳥類,以其鮮艷的羽毛和模仿人類語言的能力而聞名。鸚鵡目鳥類主要分佈在熱帶和亞熱帶地區,它們的喙和腳都經過特化,適應了啃食堅果和攀爬的生活方式。
猛禽類,包括鷹形目(Accipitriformes)和鴞形目(Strigiformes),是另一個重要的鳥類類群。這些鳥類以其銳利的喙和爪、出色的視力而著名,是生態系統中重要的頂級捕食者。鷹形目包括鷹、隼等日行性猛禽,而鴞形目則包括貓頭鷹等夜行性猛禽。
水鳥類群也展現了豐富的多樣性。鵜鶘目(Pelecaniformes)、鸛形目(Ciconiiformes)和鷺科(Ardeidae)等類群適應了濕地和水域環境,發展出了長腿和長喙等特徵。企鵝目(Sphenisciformes)則完全適應了水生生活,其翅膀演化成了游泳用的鰭狀結構。
海鳥類群,如鷗科(Laridae)和信天翁科(Diomedeidae),則適應了海洋環境。這些鳥類通常有防水的羽毛和特化的鹽腺,能夠在海水中生存。信天翁以其巨大的翼展和長距離飛行能力而聞名。
除了這些主要類群,還有許多特化的小類群,如蜂鳥科(Trochilidae),它們是世界上最小的鳥類,能夠懸停飛行;啄木鳥科(Picidae),它們的喙和舌頭特化用於從樹皮下取食昆蟲;鴕鳥科(Struthionidae),它們是現存最大的鳥類,完全失去了飛行能力。
鳥類的多樣性不僅體現在形態上,也體現在它們的行為和生態位上。例如,在繁殖行為方面,有些鳥類形成終身配對,如天鵝;有些則每年更換配偶,如許多雀形目鳥類。在築巢方面,也有巨大的差異,從簡單的地上凹坑到複雜的懸掛式巢穴,反映了不同鳥類對環境的適應。
遷徙行為是另一個展現鳥類多樣性的方面。有些鳥類,如北極燕鷗,每年進行長達數萬公里的遷徙;而另一些鳥類則是留鳥,終年不離開其棲息地。這種行為上的差異反映了鳥類對季節性資源變化的不同適應策略。
鳥類的食性也展現了極大的多樣性。有些鳥類是專性食蟲者,如食蟲目(Coraciiformes)中的翠鳥;有些是專性食果者,如極樂鳥科(Paradisaeidae);還有些是雜食性的,如烏鴉科(Corvidae)。這種食性的多樣性使得鳥類能夠佔據生態系統中的各種生態位。
值得注意的是,鳥類的分類和多樣性理解是一個動態的過程。隨著新技術的應用,特別是基因組學研究的深入,鳥類的分類系統正在不斷更新。例如,近年來的研究顯示,傳統上被歸類為鷺科的鹮鶴,實際上與朱鷺和鷸科鳥類更近緣。這種分類上的變動反映了我們對鳥類演化歷史理解的不斷深化。
鳥類的多樣性不僅體現在現存物種上,化石記錄也揭示了許多已滅絕的鳥類類群。例如,恐鳥(Phorusrhacidae)是一群已滅絕的大型肉食鳥類,曾在南美洲的生態系統中扮演重要角色。研究這些已滅絕的類群有助於我們更全面地理解鳥類的演化歷史和多樣性。
鳥類的分類和多樣性研究不僅具有重要的科學意義,也對生物多樣性保護具有重要的實際意義。通過瞭解鳥類的分類和多樣性,我們可以更好地評估生態系統的健康狀況,制定更有效的保護策略。例如,識別出的特有種或瀕危種常常成為保護工作的重點對象。
鳥類的多樣性還為我們提供了豐富的研究素材,從行為生態學到進化生物學,從生物地理學到保護生物學,鳥類都是重要的研究對象。通過研究鳥類的多樣性,我們不僅能夠更好地理解生物演化的過程,也能夠獲得關於生態系統功能和環境變化的重要資訊。
第三章 鳥類多樣性與分類
3.1 鳥類的形態特徵
鳥類是一個獨特而多樣的動物類群,其形態特徵是長期演化的結果,不僅反映了它們的生活方式和生態位,也是它們成功適應各種環境的關鍵。讓我們深入探討鳥類的主要形態特徵,以及這些特徵如何支持它們的生存和繁衍。
首先,最顯著的鳥類特徵是羽毛。羽毛是鳥類特有的表皮衍生物,由角蛋白構成,具有多重功能。羽毛不僅提供了保溫和防水的作用,還在飛行中扮演著關鍵角色。鳥類的羽毛可以分為幾種主要類型:覆羽、飛羽和絨羽。覆羽覆蓋鳥體大部分表面,提供保護和保溫;飛羽包括初級飛羽和次級飛羽,是實現飛行的主要結構;絨羽則主要起到保溫作用。不同鳥類的羽毛在結構、顏色和排列上都有很大差異,這些差異往往反映了它們的生活環境和行為習慣。
鳥類的骨骼系統是另一個重要的形態特徵。為了適應飛行,鳥類的骨骼經歷了顯著的演化。鳥類的骨骼通常較為輕盈,許多骨頭是中空的,這種結構在保持強度的同時大大減輕了重量。鳥類的頭骨高度特化,形成了輕巧而堅固的結構。頭骨與脊柱的連接方式使得鳥類能夠靈活地轉動頭部,這對於飛行和捕食都非常重要。
鳥類的脊柱也經過了特化。頸椎數量增多,使得頸部更加靈活;胸椎、腰椎和薦椎融合形成糖骨,為強大的飛行肌肉提供了堅固的附著點。尾椎則縮短並融合成尾綱骨,支撐著尾羽。鳥類的肋骨與胸骨相連,形成了堅固的胸廓,保護內臟器官並支援飛行肌肉。
鳥類的前肢演化成翅膀是其最顯著的特徵之一。翅膀由變形的手和前臂骨骼構成,覆蓋著特化的飛羽。翅膀的形狀和大小因應不同鳥類的飛行方式而有所不同。例如,海鳥通常有細長的翅膀,適合長時間滑翔;而森林鳥類則可能有較短而圓的翅膀,有利於在樹木間靈活飛行。
鳥類的後肢也經過了特化。大多數鳥類的後肢適應了站立、行走或抓握。腿骨通常較短而強壯,腳趾的數量和排列方式因應不同的生活方式而變化。例如,攀爬鳥類通常有兩趾向前、兩趾向後的排列,有利於抓握樹枝;而涉水鳥類可能有蹼狀的腳,適合在水中游動。
鳥類的喙是另一個高度特化的結構。喙由角質層覆蓋的上下頜骨構成,取代了牙齒的功能。喙的形狀和大小因應不同的食性而有極大的變化。例如,食果鳥類可能有短而寬的喙,適合捏取果實;而食魚鳥類則可能有細長的喙,有利於捕捉魚類。鴨科鳥類的喙邊緣有濾食板,用於過濾水中的食物顆粒。猛禽的喙則通常彎曲而鋒利,適合撕裂獵物。
鳥類的感官系統也有獨特的形態特徵。鳥類的眼睛通常較大,視力敏銳,這是適應飛行生活的結果。很多鳥類具有四種視錐細胞,使它們能夠感知紫外光,這在尋找食物和識別同類時非常有用。鳥類的聽覺系統也非常發達,內耳結構複雜,能夠精確定位聲音來源。某些夜行鳥類,如貓頭鷹,有特化的耳朵結構,能夠在黑暗中精確定位獵物。
鳥類的呼吸系統是其另一個獨特的形態特徵。除了肺部,鳥類還有一系列氣囊,這些氣囊與肺部和部分中空骨骼相連。這種獨特的呼吸系統使得鳥類能夠進行連續的單向氣流呼吸,大大提高了氧氣利用效率,這對於高強度的飛行活動至關重要。
鳥類的消化系統也有其特點。由於飛行需要減輕體重,鳥類沒有牙齒,而是發展出了特化的消化器官。很多鳥類有嗉囊,用於暫時儲存食物。鳥類的胃通常分為腺胃和肌胃兩部分,肌胃壁厚而有力,能夠幫助研磨食物。某些食肉鳥類能夠將無法消化的食物殘渣,如骨頭和羽毛,以鳥餌的形式吐出。
鳥類的泄殖腔是另一個重要的形態特徵。泄殖腔是消化、排泄和生殖系統的共同出口。這種結構不僅簡化了鳥類的身體構造,也減輕了體重,有利於飛行。大多數鳥類通過泄殖腔進行交配,只有少數鳥類(如鴨和鵝)保留了原始的交配器。
鳥類的體溫調節系統也值得一提。作為恆溫動物,鳥類通常保持較高的體溫(約40°C)。它們的皮膚缺乏汗腺,主要通過呼吸來散熱。一些鳥類,如禿鷲,有特化的裸露皮膚區域,可以通過改變血流來調節體溫。
鳥類的形態特徵還包括一些內部結構。例如,鳥類的心臟相對體型來說較大,這是為了滿足飛行時的高能量需求。鳥類的大腦雖然相對較小,但某些區域,如負責學習和記憶的前腦,卻相當發達。
值得注意的是,雖然這些是鳥類的普遍特徵,但在不同種類的鳥類中,這些特徵可能有顯著的變異。例如,企鵝雖然屬於鳥類,但其翅膀已經演化成了類似鰭的結構,適應了水下游泳的需求。而鴕鳥等不會飛行的鳥類,其骨骼結構和肌肉分佈與會飛的鳥類有很大不同。
鳥類的形態特徵是長期演化的結果,反映了它們對不同生態環境的適應。這些特徵不僅使鳥類成為了成功的動物類群,能夠適應從極地到熱帶、從海洋到沙漠的各種環境,也使得鳥類成為了生態系統中重要的一員,在授粉、種子傳播、害蟲控制等方面扮演著關鍵角色。
研究鳥類的形態特徵不僅有助於我們理解鳥類的生物學,也為我們提供了理解演化過程的重要視窗。通過比較不同鳥類的形態特徵,我們可以追溯它們的演化歷史,理解形態和功能之間的關係,以及環境對物種演化的影響。這些知識不僅在生物學研究中有重要意義,在生物多樣性保護、生物仿生技術等領域也有廣泛的應用前景。
3.2 鳥類多樣性的全球格局
鳥類多樣性的全球格局是一個複雜而引人入勝的主題,反映了地球上生物多樣性分佈的整體趨勢。這種格局是地理、氣候、生態和進化因素長期相互作用的結果,展現了鳥類如何適應和繁衍在地球的各個角落。
首先,我們需要瞭解鳥類多樣性在全球的整體分佈趨勢。一般而言,鳥類物種豐富度從赤道向兩極遞減,這種現象被稱為緯度多樣性梯度。熱帶地區,特別是熱帶雨林,擁有最豐富的鳥類多樣性。例如,哥倫比亞、秘魯和巴西等南美國家擁有世界上最多的鳥物種。這種分佈格局可以從幾個方面來解釋。
首先,熱帶地區全年穩定的高溫和充足的降水為鳥類提供了豐富的食物資源和多樣的棲息地。穩定的氣候條件也允許物種進行更精細的生態位分化,從而支持更多物種共存。相比之下,溫帶和寒帶地區的季節性變化更為劇烈,限制了能夠全年生存的物種數量。
其次,熱帶地區的進化時間較長。地質歷史上,熱帶地區受冰川作用的影響較小,提供了相對穩定的環境,使得物種有更長的時間進行適應性輻射。這導致了更高的特有種比例和整體多樣性。
地形複雜性是影響鳥類多樣性的另一個重要因素。山區往往擁有較高的鳥類多樣性,這是因為山地環境提供了多樣的小氣候和棲息地類型。安第斯山脈、喜馬拉雅山脈等大型山系是鳥類多樣性的熱點。山區的隔離效應也促進了物種形成,導致了高度的地方特有性。
島嶼生物地理學在解釋鳥類多樣性格局中也扮演著重要角色。大型島嶼通常比小型島嶼擁有更多的鳥物種,這與島嶼生物地理學理論相符。同時,島嶼的隔離程度也影響著其鳥類多樣性。較為隔離的島嶼往往擁有較高比例的特有種,如加拉帕戈斯群島和夏威夷群島。
歷史因素,特別是板塊運動和古氣候變化,對當前的鳥類多樣性格局產生了深遠影響。例如,南美洲長期的地理隔離導致了該地區獨特而豐富的鳥類區系。澳大利亞由於長期隔離,發展出了特有的鳥類群,如鸚鵡目和食火雞目的多樣化。
生態系統的類型和複雜性直接影響著鳥類的多樣性。熱帶雨林由於其複雜的垂直結構和豐富的植物多樣性,為鳥類提供了多樣的生態位。相比之下,沙漠和極地苔原等環境條件苛刻的生態系統,鳥類多樣性相對較低,但往往擁有高度特化的物種。
水體的存在也是影響鳥類多樣性的重要因素。河流、湖泊和濕地往往是鳥類多樣性的熱點。這些水體不僅為水鳥提供了棲息地,也為陸地鳥類提供了水源和豐富的食物資源。沿海地區,特別是紅樹林和河口區,往往擁有豐富的鳥類多樣性。
遷徙模式也塑造著全球鳥類多樣性格局。許多鳥物種在不同季節在南北半球之間進行長距離遷徙。這種行為使得某些地區的鳥類多樣性呈現季節性變化。例如,北美和歐洲的溫帶地區在夏季時鳥類多樣性顯著增加,這是由於大量候鳥的到來。
人類活動對鳥類多樣性格局產生了深遠影響。棲息地破壞、過度捕獵和氣候變化等因素導致了許多地區鳥類多樣性的下降。特別是在熱帶地區,森林砍伐對鳥類多樣性造成了嚴重威脅。相反,一些人為環境,如城市公園和農田,為某些適應能力強的鳥物種提供了新的棲息地,改變了局部的多樣性格局。
在研究鳥類多樣性的全球格局時,不同的多樣性指標可能會得出不同的結論。除了物種豐富度,我們還需要考慮功能多樣性和系統發育多樣性。功能多樣性反映了鳥類在生態系統中所扮演的不同角色,而系統發育多樣性則反映了進化歷史的深度。某些地區可能物種數量不多,但擁有獨特的功能群或古老的譜系,因此在保護生物多樣性方面具有特殊重要性。
全球鳥類多樣性熱點往往與其他生物類群的多樣性熱點重疊,這反映了影響生物多樣性的一些共同因素。然而,鳥類由於其獨特的飛行能力和適應性,其多樣性格局也展現出一些特有的特徵。例如,某些海洋鳥類可以在貧瘠的海洋環境中形成高度多樣化的群落。
鳥類多樣性的全球格局並非靜態的,而是隨時間動態變化的。長期的氣候變化、生態系統演替和人類活動等因素都在持續塑造著這一格局。例如,全球變暖正在改變許多鳥物種的分佈範圍,導致某些地區的鳥類群落組成發生變化。
瞭解鳥類多樣性的全球格局對於制定有效的保護策略至關重要。通過識別多樣性熱點和特有種中心,我們可以優先保護那些對全球鳥類多樣性貢獻最大的地區。同時,鳥類作為良好的環境指示物種,其多樣性格局的變化可以反映更廣泛的生態系統健康狀況。
鳥類多樣性的全球格局研究不僅有助於我們理解當前的生物多樣性分佈,也為預測未來的變化提供了基礎。隨著全球環境的快速變化,持續監測和研究這些格局變得越來越重要。只有深入理解影響鳥類多樣性分佈的各種因素,我們才能更好地保護這些美麗而重要的生物,維護地球的生態平衡。
3.3 鳥類的適應性輻射
鳥類的適應性輻射是進化生物學中一個引人入勝的主題,展示了生物如何通過自然選擇在相對短的時間內快速分化,適應不同的生態位。這個過程不僅塑造了鳥類的驚人多樣性,也為我們理解生物進化的機制提供了寶貴的洞察。
適應性輻射通常發生在生態機會出現時,比如新的棲息地變得可用,或者競爭者消失。在鳥類的演化歷史中,我們可以看到多次顯著的適應性輻射事件,每次都產生了一系列適應不同生態位的新種類。
其中最著名的例子之一是達爾文雀的適應性輻射。這群鳥類在加拉帕戈斯群島上的演化過程為達爾文提供了關鍵的進化證據。原本來自南美大陸的祖先種抵達這些火山島後,面對各種不同的食物資源和生態環境,迅速分化成了多個不同的種類。這些種類主要在喙部形態上表現出顯著差異,反映了它們對不同食物資源的適應。有的發展出強壯的喙部用於破開堅果,有的則演化出細長的喙部以提取花蜜。這個例子生動地展示了如何通過適應性輻射填補不同的生態位。
另一個引人注目的適應性輻射例子是夏威夷蜜鳥。這個家族從一個祖先種演化出超過50個種類,展現了令人驚嘆的形態和行為多樣性。有些種類發展出彎曲的喙部以適應特定植物的花形,有些則演化出直喙以捕食昆蟲。這種多樣化不僅體現在喙部形態上,還包括羽色、體型和行為習性等多個方面。夏威夷蜜鳥的適應性輻射展示了島嶼環境如何促進快速的物種形成和生態適應。
在南美洲,探鳥類提供了另一個精彩的適應性輻射案例。這個家族包含了約330個種類,從小巧的蜂鳥到大型的巨嘴鳥,展現了驚人的形態多樣性。探鳥類的適應性輻射涵蓋了多種生態位,包括取食昆蟲、水果、花蜜,甚至小型脊椎動物。這個例子說明瞭如何通過適應不同的食物資源和取食策略來實現生態分化。
非洲大陸上,奇異鳥科的適應性輻射也值得關注。這個家族在不同的森林和草原棲息地中演化出多種形態,適應各種不同的生態位。有些種類專門在地面覓食,有些則更喜歡在樹冠層活動。這種多樣化反映了非洲大陸豐富多樣的生態環境。
在澳大利亞和新幾內亞地區,鸚鵡目的適應性輻射尤為顯著。從小型的虎皮鸚鵡到大型的金剛鸚鵡,鸚鵡類展現了令人驚嘆的形態和行為多樣性。它們適應了從熱帶雨林到乾旱內陸的各種棲息地,發展出多種取食策略,包括取食種子、水果、花蜜和昆蟲。鸚鵡類的適應性輻射不僅體現在形態上,還包括了複雜的社會行為和認知能力的演化。
鳥類的適應性輻射並不僅限於特定的地理區域或分類群。在全球範圍內,我們可以觀察到鳥類如何適應各種極端環境。例如,企鵝科的適應性輻射使得這些鳥類能夠在南半球的各種水生環境中繁衍,從溫帶海岸到南極洲的冰雪世界。每個企鵝種類都展現出對特定環境的獨特適應,包括體型、羽毛結構和行為方面的變化。
雁鴨科提供了另一個全球性適應性輻射的例子。這個家族的成員遍佈全球各種水生和半水生環境,從北極苔原到熱帶沼澤。它們在喙部形態、腳蹼結構和羽毛特徵等方面表現出顯著的多樣性,反映了對不同水體環境和食物資源的適應。
鳥類的適應性輻射不僅體現在形態上,還包括行為和生理特徵的演化。例如,蜂鳥科的適應性輻射不僅產生了多種喙部形態以適應不同的花朵,還涉及了飛行能力、新陳代謝和感知能力的特殊適應。蜂鳥能夠懸停飛行和倒飛的能力,以及它們獨特的能量代謝方式,都是適應性輻射過程中演化出的創新特徵。
在一些情況下,鳥類的適應性輻射可能導致趨同進化。例如,在不同大陸上,一些不相關的鳥類科屬可能演化出相似的形態和行為,以適應相似的生態位。這種現象在島嶼環境中尤為常見,如新西蘭的幾種已滅絕的巨型鳥類就在生態功能上類似於大型哺乳動物。
鳥類的適應性輻射還涉及到感官系統的演化。例如,貓頭鷹科的適應性輻射不僅涉及了適應不同獵物和棲息地的形態變化,還包括了聽覺和視覺系統的高度特化。這使得不同種類的貓頭鷹能夠在各種光照條件下有效地捕獵。
值得注意的是,鳥類的適應性輻射過程並非總是平穩進行的。環境變化、新捕食者的出現或競爭者的入侵都可能打斷或改變輻射的方向。例如,人類引入的捕食者和競爭者已經導致了許多島嶼鳥類多樣性的下降,某些情況下甚至導致了整個輻射支系的滅絕。
研究鳥類的適應性輻射不僅有助於我們理解生物多樣性的形成過程,還為保護生物學提供了重要見解。通過瞭解特定地區或生態系統中鳥類的演化歷史和適應過程,我們可以更好地預測和應對環境變化對鳥類多樣性的影響。
鳥類的適應性輻射研究還揭示了生態機會如何驅動進化創新。例如,某些鳥類群體在進入新的生態位後,可能會發展出全新的形態或行為特徵。這種創新可能進一步開啟新的適應機會,形成一種正反饋循環,推動更廣泛的多樣化。
隨著分子生物學和基因組學技術的發展,我們對鳥類適應性輻射的理解正在不斷深化。這些新技術使我們能夠探索支撐形態和行為多樣性的遺傳基礎,揭示適應性輻射過程中基因組水準的變化。這些研究不僅幫助我們理解過去的演化過程,還為預測未來的演化趨勢提供了工具。
3.4 自然選擇與配偶選擇
鳥類的演化過程是自然選擇和配偶選擇共同作用的結果,這兩種機制塑造了鳥類的多樣性,並使它們能夠適應各種環境。自然選擇主要關注生存和繁衍能力,而配偶選擇則聚焦於吸引異性和繁殖成功。這兩種選擇壓力共同推動了鳥類的演化,產生了我們今天所見的豐富多彩的鳥類世界。
自然選擇是達爾文提出的核心進化理論,它解釋了生物如何適應其環境。在鳥類中,自然選擇作用於各種特徵,包括形態、生理和行為。例如,鳥喙的形狀和大小直接受到食物類型的影響。加拉帕戈斯雀的研究就是一個經典案例,不同島嶼上的鳥喙形態差異反映了它們對當地食物資源的適應。在乾旱季節,堅果成為主要食物來源時,具有更強壯喙部的個體存活率更高,這種選擇壓力over時間推動了鳥喙形態的演化。
自然選擇還影響鳥類的體型和羽毛。例如,生活在寒冷地區的鳥類通常體型較大,這有助於減少相對表面積,從而減少熱量損失。極地的帝企鵝就是一個很好的例子,它們龐大的體型有助於在嚴寒環境中保持體溫。相反,熱帶地區的蜂鳥則體型小巧,有利於在高溫環境中散熱。羽毛的顏色和結構也受到自然選擇的影響,常常用於偽裝。例如,雷鳥的羽毛會隨季節變化顏色,在冬季變為白色以融入積雪環境,這大大提高了它們避免被捕食者發現的能力。
鳥類的行為特徵同樣受到自然選擇的塑造。遷徙行為就是一個顯著的例子。能夠成功往返於繁殖地和越冬地的鳥類更有可能存活並繁衍後代。這種選擇壓力導致了複雜的遷徙模式的演化,包括精確的導航能力和高效的飛行技巧。另一個例子是築巢行為,能夠建造安全、隱蔽巢穴的鳥類更有可能成功孵化和撫養幼鳥,這推動了多樣化的築巢技術的演化。
然而,僅靠自然選擇無法完全解釋鳥類的某些特徵,特別是那些看似不利於生存的特徵。這就是配偶選擇發揮作用的地方。配偶選擇是性選擇的一種形式,它解釋了為什麼某些特徵會演化,即使這些特徵可能降低個體的生存能力。在鳥類中,配偶選擇通常表現為雌性對雄性特徵的偏好。
孔雀的華麗尾羽是配偶選擇的經典例子。這些巨大、色彩鮮艷的羽毛實際上可能增加了被捕食的風險,並且在飛行時造成不便。然而,雌性孔雀偏好具有更大、更艷麗尾羽的雄性。這種偏好可能源於這些特徵表明了雄性的健康狀況和基因質量。over時間,這種選擇壓力導致了雄性孔雀尾羽的誇張演化。
鳥類的歌唱能力也是配偶選擇的重要對象。許多鳥類物種中,雄鳥通過複雜的歌聲來吸引雌鳥。能夠演唱更複雜、更持久歌曲的雄鳥通常更受雌鳥青睞。這種選擇壓力推動了鳥類發聲器官和大腦控制區域的演化,使得某些物種能夠產生極其複雜和多樣的歌聲。夜鶯和模仿鳥就以其出色的歌唱能力而聞名。
配偶選擇還可能導致性二態性的演化,即雌雄個體在外觀上的顯著差異。在許多鳥類物種中,雄性比雌性更具有鮮艷的羽毛或誇張的裝飾。天堂鳥就是一個極端的例子,雄鳥具有極其華麗和複雜的羽毛裝飾,而雌鳥則相對樸素。這種差異是長期配偶選擇的結果,雌鳥持續選擇最鮮艷、最引人注目的雄鳥作為配偶。
有趣的是,配偶選擇並不總是導致雄性特徵的誇張。在某些物種中,如澳大利亞的刺尾鷹,情況恰恰相反。雌鳥比雄鳥更大、更具有攻擊性,並且在求偶中扮演主動角色。這種「角色反轉」可能是由特定的生態壓力和繁殖策略推動的。
自然選擇和配偶選擇並非相互獨立的過程,它們經常相互作用,有時甚至相互對抗。例如,雖然鮮艷的羽毛可能增加吸引配偶的機會,但也可能增加被捕食者發現的風險。因此,鳥類的演化常常是這兩種選擇力量之間平衡的結果。
在某些情況下,自然選擇和配偶選擇可能產生協同效應。例如,某些鳥類的羽毛顏色不僅用於吸引配偶,還可能指示個體的健康狀況。紅色或黃色的羽毛常常由胡蘿蔔素產生,這些色素也具有增強免疫系統的功能。因此,選擇鮮艷羽毛的雌鳥可能同時選擇了更健康的配偶,這既滿足了配偶選擇的標準,也符合自然選擇的要求。
鳥類的配對系統也受到自然選擇和配偶選擇的共同影響。例如,在資源豐富、穩定的環境中,可能會演化出終身配對的系統,因為雙親共同撫育後代可以提高繁殖成功率。相反,在資源不穩定或分佈不均的環境中,可能會演化出多配或混合配對策略,以最大化繁殖成功的機會。
值得注意的是,鳥類的演化並非一個靜態的過程,而是持續的、動態的。環境變化、新的生態壓力或機會都可能改變選擇壓力的方向和強度。例如,人類活動導致的棲息地變化可能改變某些鳥類特徵的適應價值,從而影響其演化軌跡。
此外,鳥類的演化還涉及許多其他因素,如基因漂變、基因流動和突變。這些因素與自然選擇和配偶選擇相互作用,共同塑造了鳥類的遺傳結構和表型特徵。例如,小種群中的基因漂變可能導致某些特徵的快速固定,即使這些特徵並不具有明顯的適應意義。
隨著分子生物學和基因組學技術的發展,我們對鳥類演化的理解正在不斷深化。這些新技術使我們能夠探索支撐形態和行為多樣性的遺傳基礎,揭示自然選擇和配偶選擇在基因組水準上的作用機制。這些研究不僅幫助我們理解過去的演化過程,還為預測未來的演化趨勢提供了工具。
研究鳥類的演化過程不僅有助於我們理解生物多樣性的形成機制,還為保護生物學提供了重要見解。通過瞭解自然選擇和配偶選擇如何塑造鳥類特徵,我們可以更好地預測和應對環境變化對鳥類物種的影響,從而制定更有效的保護策略。
物種形成與雜交 How Birds Evolve Speciation and Hybridization
物種形成和雜交是鳥類演化過程中兩個密切相關且極為重要的現象。物種形成是新物種產生的過程,而雜交則是不同物種或亞種之間基因交流的途徑。這兩個過程在鳥類的演化歷史中扮演著關鍵角色,共同塑造了我們今天所見的鳥類多樣性。
物種形成通常被認為是演化的核心過程之一。在鳥類中,物種形成可以通過多種機制發生,其中最常見的是異域物種形成和同域物種形成。異域物種形成發生在地理上分隔的種群之間,而同域物種形成則發生在共用同一地理區域的種群之間。
異域物種形成在鳥類中尤為常見。當一個物種的不同種群被地理屏障(如山脈、海洋或沙漠)分隔時,它們可能會逐漸演化出不同的特徵。例如,加拉帕戈斯群島上的達爾文雀就是異域物種形成的經典案例。這些鳥類的祖先可能來自南美大陸,在抵達不同的島嶼後,面對不同的食物資源和環境條件,逐漸演化出適應當地生態位的特徵,最終形成了多個不同的物種。
島嶼環境特別有利於異域物種形成。夏威夷蜜鳥科的輻射適應就是另一個引人注目的例子。這個家族從一個祖先物種演化出約50個物種,每個物種都適應了特定島嶼或生態位的環境。這種快速的物種形成過程展示了地理隔離如何促進生物多樣性的產生。
同域物種形成雖然在鳥類中較為罕見,但也確實存在。這種物種形成通常涉及生態位分化或行為隔離。例如,非洲的灰胸鸚鵡存在兩種生態型,一種主要在森林中生活,另一種主要在草原生活。儘管它們地理上重疊,但由於棲息地偏好的差異,它們很少交配,可能正在經歷同域物種形成的過程。
在物種形成過程中,生殖隔離機制的演化至關重要。這些機制可以是前合子障礙(如行為、生態或機械隔離)或後合子障礙(如雜種不育或不活)。在鳥類中,行為隔離尤為重要,特別是通過求偶展示和鳴叫的差異來實現。例如,近緣的鳥類種可能有非常相似的外表,但它們的鳴叫可能完全不同,這有效地防止了它們之間的交配。
然而,物種形成並非總是一個完整或不可逆的過程。這就是雜交在鳥類演化中發揮作用的地方。雜交是指不同物種或亞種之間的交配,它可以導致基因流動,有時甚至可以逆轉物種形成過程或促進新物種的形成。
雜交在鳥類中相對常見,估計約有10%的鳥類物種會與近緣種發生雜交。雜交可能發生在物種形成的各個階段,從剛開始分化的種群到已經明顯分化的物種之間。雜交的結果可能有多種:它可能產生不育的後代,從而加強物種間的生殖隔離;也可能產生可育的後代,導致基因滲入或甚至形成新的混合種。
基因滲入是雜交的一個重要結果,指的是一個物種的基因通過雜交進入另一個物種的基因庫。這種現象可能導致適應性狀的轉移,增加遺傳變異,甚至模糊物種界限。例如,北美的黃嘴地鶇和灰嘴地鶇之間的雜交導致了廣泛的基因滲入,使得這兩個物種的遺傳組成變得更加相似。
在某些情況下,雜交可能導致混合種的形成。義大利斑斯巴木鶯就是一個著名的例子。這個物種被認為是由歐洲斑斯巴木鶯和撒丁斑斯巴木鶯雜交形成的。混合種的形成展示了雜交如何創造新的遺傳組合,potential leading to新物種的產生。
雜交還可能在適應性進化中發揮重要作用。通過雜交,有利的基因變異可以在不同物種之間傳播,potential accelerating適應過程。例如,研究表明,家雞的黃皮膚特徵可能源於與灰原雞的雜交,這種特徵隨後被人類在家雞育種過程中選擇和固定。
然而,雜交也可能帶來負面影響。在某些情況下,大規模的雜交可能威脅到稀有物種的遺傳完整性。例如,美國的金翅雀正面臨著與引入的歐洲金翅雀雜交的威脅,這可能導致本地物種的基因庫被稀釋。
氣候變化和棲息地破壞等人為因素正在改變許多鳥類物種的分佈範圍,增加了以前不常接觸的物種之間雜交的機會。這種情況可能加速物種形成過程,也可能威脅現有物種的完整性。例如,由於全球變暖,北極的白頰黑雁正與加拿大黑雁的棲息地重疊增加,導致這兩個物種之間的雜交頻率上升。
研究物種形成和雜交對於理解鳥類的演化動態至關重要。這些過程不僅揭示了生物多樣性如何產生和維持,還為我們理解物種概念本身提供了洞見。傳統上,物種被定義為能夠相互交配並產生可育後代的個體群體。然而,鳥類中普遍存在的雜交現象挑戰了這一定義,表明物種界限可能比我們想像的更加模糊和動態。
近年來,基因組學技術的發展為研究物種形成和雜交提供了新的工具。通過分析全基因組數據,科學家們能夠重建物種的演化歷史,識別雜交事件,並追蹤特定基因或適應性狀在物種之間的傳播。這些研究不僅加深了我們對鳥類演化過程的理解,還為保護生物學提供了重要指導。
例如,基因組分析揭示了許多被認為是單一物種的鳥類實際上可能包含多個隱蔽種。這些發現對保護工作有重要影響,因為它們可能改變我們對物種多樣性和保護優先級的認識。另一方面,基因組研究也顯示了一些形態上截然不同的鳥類之間存在基因流動,這突顯了保護整個生態系統而不僅僅是單個物種的重要性。
物種形成和雜交的研究還為應對氣候變化和其他人為威脅提供了見解。通過瞭解這些過程如何影響鳥類的適應能力和遺傳多樣性,我們可以更好地預測和管理人類活動對鳥類種群的影響。例如,在某些情況下,促進不同種群之間的基因流動可能有助於增加瀕危物種的遺傳多樣性和適應潛力。
3.5 鳥類的分類體系
鳥類是一個多樣化且引人入勝的動物類群,其分類和命名體系反映了科學家對這些生物的深入研究和理解。鳥類屬於脊椎動物門(Vertebrata)中的鳥綱(Aves),是恐龍演化而來的後裔。現代鳥類分類主要基於形態特徵、行為和遺傳學證據,隨著分子生物學技術的發展,DNA分析在鳥類分類中扮演著越來越重要的角色。
鳥類的分類遵循林奈分類系統,從綱到種依次為:綱、目、科、屬、種。目前,全球已知的鳥類物種約有10,000多種,分為約40個目。其中最大的目是雀形目(Passeriformes),包括了超過半數的鳥類物種,如麻雀、燕子和烏鴉等。其他常見的目包括鸚鵡目(Psittaciformes)、鴿形目(Columbiformes)和鵜形目(Pelecaniformes)等。
在科級分類中,鳥類被劃分為約230個科。每個科通常包含多個屬,而每個屬又包含一個或多個種。例如,鴨科(Anatidae)包括了鴨、鵝和天鵝等水禽;鶯科(Sylviidae)則包括了多種小型鳴禽。
鳥類的命名遵循國際動物命名法規(ICZN),採用二名法。每個物種都有一個由兩部分組成的拉丁學名:屬名和種加詞。屬名總是大寫,而種加詞則小寫。例如,家鴿的學名是Columba livia,其中Columba是屬名,livia是種加詞。此外,鳥類還有通用名,即我們日常使用的俗名,如「麻雀」、「鴿子」等。
鳥類的分類和命名是一個動態的過程,隨著新證據的出現和研究方法的進步而不斷更新。近年來,分子系統學的發展對鳥類分類產生了重大影響。通過比較不同物種的DNA序列,科學家們能夠更準確地確定物種間的親緣關係,有時甚至導致了分類體系的重大調整。
例如,鴕鳥、鴯鶓和幾維鳥等不會飛的鳥類曾被歸類為「走禽」,與「飛禽」相對。然而,基因研究表明,這些不會飛的鳥類並不是一個單系群,而是在演化過程中多次獨立喪失了飛行能力。因此,現代分類系統不再使用「走禽」和「飛禽」這樣的分類。
另一個顯著的例子是鸚鵡目和鳩鴿目之間的關係。傳統上,這兩個目被認為是相距較遠的類群,但分子證據顯示它們實際上是親緣關係較近的姐妹群。這種發現促使分類學家重新考慮了這些鳥類的演化歷史和分類位置。
鳥類的命名也反映了科學發現和文化背景。許多鳥類的學名源自拉丁語或古希臘語,描述了鳥類的外觀、行為或棲息地。例如,烏鴉屬的學名Corvus來自拉丁語,意為「烏鴉」;而夜鷹科的學名Caprimulgidae則源自一個古老的誤解,意為「擠山羊奶的人」,因為人們曾錯誤地認為這些鳥會在夜間吸食山羊的奶。
同時,鳥類的通用名往往反映了當地文化和語言的特點。例如,同一種鳥在不同國家可能有完全不同的名字。這種多樣性既豐富了鳥類學的文化內涵,也為國際交流帶來了挑戰,因此科學界更加依賴統一的學名系統。
隨著對鳥類認知的深入,分類系統也在不斷細化。亞種的概念在鳥類分類中扮演著重要角色,用來描述同一物種內的地理變異。亞種通常用三名法表示,在物種學名後加上亞種名。例如,歐亞大陸的家鴿(Columba livia livia)和北美洲的家鴿(Columba livia domestica)就是兩個不同的亞種。
鳥類的分類和命名不僅是學術研究的基礎,也對鳥類保護和管理具有重要意義。準確的分類有助於評估物種的保護狀況,制定合適的保護策略。例如,當一個廣泛分佈的物種被分為多個獨立的物種時,每個新物種的分佈範圍和種群數量都會相應減少,可能需要更緊急的保護措施。
此外,鳥類分類還影響著生物地理學、生態學和進化生物學等相關領域的研究。通過研究不同鳥類類群的分佈模式和演化歷史,科學家們可以更好地理解地球生物多樣性的形成過程和未來變化趨勢。
鳥類學家們持續致力於完善分類系統,解決分類爭議。例如,關於某些近緣物種是否應該被視為獨立物種或亞種,常常存在爭議。這些爭議的解決不僅依賴於形態學和遺傳學證據,還需要考慮生態位元、行為和繁殖隔離等因素。
在鳥類分類和命名的未來發展中,整合形態學、行為學、生態學和基因組學等多學科方法將變得越來越重要。新技術如高通量測序和生物資訊學分析,將為鳥類系統發育研究提供更多精確的數據。同時,鳥類學家們也在努力建立全球性的鳥類分類數據庫,以促進國際合作和知識共用。
總之,鳥類的分類和命名是一個複雜而動態的領域,反映了人類對這些迷人生物的持續探索和理解。隨著科技的進步和研究方法的創新,我們對鳥類多樣性的認識將不斷深化,為生物多樣性保護和生態系統管理提供更堅實的科學基礎。
3.6 鳥類種群分化的驅動力
鳥類種群分化是一個複雜而持續的過程,由多種因素共同驅動。這些驅動力塑造了鳥類的演化軌跡,最終導致了我們今天所見的豐富多樣的鳥類世界。理解這些驅動力不僅有助於我們解釋現有的鳥類多樣性,還能幫助我們預測未來的演化趨勢和制定有效的保護策略。
地理隔離是鳥類種群分化的最主要驅動力之一。當一個物種的不同種群被山脈、海洋、沙漠等地理屏障分隔時,它們之間的基因交流被阻斷,各自在不同的環境中演化。這種情況下,即使最初的差異很小,隨著時間的推移,這些種群可能會逐漸演化出顯著的差異。例如,大洋島嶼上的鳥類常常展現出明顯的種群分化。夏威夷群島上的蜜鳥科鳥類就是一個典型案例,原本來自同一祖先的種群在不同島嶼上演化出了多樣的形態和行為特徵。
環境異質性是另一個重要的驅動力。即使在沒有明顯地理屏障的情況下,不同的環境條件也可能導致種群分化。這包括氣候差異、食物資源的變化、捕食壓力的不同等。例如,北美大陸的家燕種群在東西兩岸表現出微妙但顯著的差異,這可能與兩地不同的氣候條件有關。環境異質性驅動的分化通常是漸進的,可能導致沿環境梯度的漸變式變異。
選擇壓力是推動鳥類種群分化的核心機制。不同環境中的選擇壓力可能偏好不同的性狀,導致種群在形態、生理或行為上的分化。例如,達爾文雀的喙部形態在不同的加拉帕戈斯群島上表現出顯著差異,這反映了各島嶼上不同食物資源的選擇壓力。自然選擇可以迅速改變種群的特徵,特別是在面臨新環境或急劇變化的情況下。
性選擇也是種群分化的重要驅動力。鳥類中的求偶行為、羽色和鳴叫等性狀常常受到強烈的性選擇。如果不同種群中雌性的偏好發生變化,可能會導致雄性性狀的快速分化。例如,天堂鳥科的不同物種展現出極其多樣的羽飾和求偶舞蹈,這很大程度上是由性選擇驅動的。性選擇可以加速種群分化,有時甚至可能導致生殖隔離的形成。
基因漂變在小種群中可能成為重要的分化驅動力。當種群規模較小時,隨機的基因頻率變化可能導致某些特徵在種群中固定或消失,即使這些特徵並不具有明顯的適應意義。這種效應在島嶼或隔離棲息地的鳥類種群中特別明顯。例如,一些小島上的鳥類可能表現出與大陸種群明顯不同的特徵,部分原因可能就是基因漂變的結果。
遷徙行為的變化也可能導致種群分化。鳥類的遷徙路線和時間通常是遺傳決定的。如果部分種群改變了遷徙模式,可能會導致與原有種群的隔離,進而促進分化。例如,歐洲的歌鶇存在遷徙和留居兩種行為類型,這可能是種群分化的早期階段。
人類活動正日益成為鳥類種群分化的重要驅動力。棲息地破碎化可能隔離原本連續分佈的種群,而城市化則可能為某些鳥類創造新的選擇壓力。例如,城市中的烏鶇相比其鄉村同類,傾向於更早開始晨鳴,這可能是對城市噪音的適應。氣候變化也正在改變許多鳥類的分佈範圍和生活週期,potential creating 新的分化機會。
入侵種的引入可能對本地鳥類種群產生深遠影響,有時會導致快速的演化響應。例如,夏威夷的一些本地鳥類在面對引入的瘧蚊時,正在演化出對禽瘧的抗性。這種情況下,不同種群面臨的選擇壓力差異可能導致顯著的分化。
棲息地特化是另一個可能導致種群分化的因素。當不同的種群適應特定的棲息地類型時,它們可能逐漸演化出不同的形態或行為特徵。例如,同一鳥種在森林和草原棲息地中的種群可能在羽色、體型或鳴叫上表現出差異。這種分化如果持續下去,最終可能導致生態種的形成。
食性的改變也可能驅動種群分化。當不同種群開始利用不同的食物資源時,它們可能演化出適應這些資源的特殊特徵。喙部形態的差異常常反映了這種分化。例如,夏威夷蜜鳥科的不同物種演化出了適應不同花型的各種喙部形態。
行為的變化可能是種群分化的另一個驅動因素。新的行為模式,如新的覓食策略或築巢方式,可能導致種群在生態上的分化。例如,歐亞大陸的烏鴉存在使用工具和不使用工具的種群,這種行為差異可能最終導致更廣泛的分化。
寄生蟲和疾病也可能推動種群分化。不同地區的種群可能面臨不同的寄生蟲壓力,從而演化出不同的免疫應對。這可能導致種群在生理特徵上的分化,甚至影響到配偶選擇,因為健康狀況通常是重要的選擇標準。
鳥類種群的分化常常是多種驅動力共同作用的結果。例如,地理隔離可能首先分開種群,然後環境差異和選擇壓力進一步推動分化。在某些情況下,初始的小幅分化可能通過正反饋機制被放大,最終導致顯著的種群差異。
理解這些驅動種群分化的力量對於鳥類保護具有重要意義。例如,認識到環境異質性對維持遺傳多樣性的重要性,可以幫助我們設計更有效的保護區網絡。同樣,瞭解人類活動如何影響鳥類演化,可以幫助我們預測和減緩人為因素對鳥類多樣性的負面影響。
隨著基因組學技術的發展,我們對鳥類種群分化的遺傳基礎有了更深入的理解。全基因組分析使我們能夠識別與適應性狀相關的基因變異,追蹤這些變異在不同種群間的擴散和分化。這些研究不僅揭示了分化的遺傳機制,還為我們理解演化速率和預測未來變化提供了新的視角。
鳥類種群分化的研究還強調了保護演化潛力的重要性。僅僅保護當前的物種多樣性是不夠的,我們還需要維護能夠產生未來多樣性的生態和演化過程。這意味著保護足夠大的種群規模、維持種群間的連通性、以及保護多樣的棲息地類型,以確保鳥類能夠繼續響應不斷變化的環境條件。
3.7 台灣 常見鳥目與 科
雞形目
雞形目(Galliformes)是鳥類分類中一個重要的目,其命名源自拉丁語"gallus",意為「公雞」。這個名稱反映了家雞在這個鳥類群體中的代表性地位。雞形目包括了許多經濟和文化上極為重要的鳥類,如雞、火雞、孔雀、鵪鶉等。這個目的命名突顯了家雞作為人類馴化最早、分佈最廣的家禽之一的重要性。
雞形目鳥類具有一系列獨特的特徵,使它們在鳥類世界中易於識別。首先,這些鳥類通常體型較大,身軀豐滿,有強壯的腿部和爪子,適合在地面行走和刨土覓食。它們的翅膀相對較短而圓,雖然能夠短距離飛行,但大多數時間都在地面活動。這種身體結構反映了它們主要的地棲生活方式。
雞形目鳥類的喙部結構是另一個顯著特徵。它們通常有短而彎曲的喙,堅固而有力,適合啄食各種食物,包括種子、漿果、嫩葉和小型動物。這種多樣化的食性使得雞形目鳥類能夠適應各種環境。
在頭部特徵上,許多雞形目鳥類都有獨特的裝飾物,如冠、垂肉或其他肉質附屬物。這些特徵在求偶展示和種內識別中扮演重要角色。例如,火雞的特徵性肉垂和孔雀華麗的尾羽都是著名的例子。
雞形目鳥類的繁殖行為也有其特點。大多數物種實行一夫多妻制,雄性通常有鮮艷的羽毛和誇張的求偶展示行為。相比之下,雌性的羽色往往較為樸素,這有助於在孵卵和育雛期間的保護色。
在生態適應方面,雞形目鳥類展現出驚人的多樣性。雖然許多物種偏好森林和灌木叢棲息地,但也有適應高山、草原甚至沙漠環境的物種。例如,雪雞能夠在高海拔的嚴酷環境中生存,而沙雞則適應了乾旱地區的生活。
雞形目鳥類在人類文化和經濟中佔有重要地位。家雞是全球最普遍的家禽,提供了大量的肉類和蛋類產品。火雞在北美洲的感恩節文化中具有象徵意義。此外,許多雞形目鳥類,如孔雀和錦雞,因其美麗的羽毛而在觀賞和文化領域中備受青睞。
然而,由於棲息地破壞和過度捕獵,一些野生雞形目鳥類面臨著生存威脅。例如,某些雉類物種已經瀕臨絕種。這凸顯了保護這些鳥類及其棲息地的重要性,不僅對於維護生物多樣性,也對於保存人類文化遺產至關重要。
總的來說,雞形目鳥類以其獨特的形態特徵、多樣的生態適應能力以及與人類文明的密切聯繫而著稱。它們不僅是重要的經濟資源,也是生態系統中不可或缺的成員,在全球範圍內的自然和文化景觀中扮演著重要角色。
鴕鳥目
鴕鳥目(Struthioniformes)是鳥類分類中一個獨特而引人注目的群體,其命名源自拉丁語"struthio",意為「鴕鳥」。這個名稱直接反映了鴕鳥作為該目中最具代表性和最為人所知的成員。鴕鳥目的命名突顯了這些大型、不會飛行的鳥類在演化史上的特殊地位。
鴕鳥目鳥類最顯著的特徵是它們完全喪失了飛行能力。這一特徵是其演化過程中的重要適應,使得它們在地面生活方式中獲得了獨特的優勢。隨著飛行能力的喪失,鴕鳥目鳥類發展出了一系列適應地面生活的特徵。
首先,鴕鳥目鳥類以其巨大的體型而聞名。鴕鳥是現存鳥類中最大的,成年鴕鳥可以達到2.7米高,重達150公斤。這種巨大的體型使它們幾乎沒有天敵,同時也讓它們能夠在開闊的草原上遠距離巡視周圍環境。
鴕鳥目鳥類的腿部極為發達,這是它們適應快速奔跑的結果。它們的腿骨粗壯而長,肌肉發達,使它們能夠達到驚人的奔跑速度。鴕鳥可以保持每小時60公里的速度持續奔跑,這使它們成為陸地上最快的兩足動物之一。
與其他鳥類不同,鴕鳥目鳥類的腳部結構也有顯著特點。它們通常只有兩個或三個腳趾,而不是典型鳥類的四趾。這種腳部結構增強了它們在奔跑時的穩定性和效率。例如,鴕鳥只有兩個腳趾,其中一個特別發達,能夠承受大部分體重。
鴕鳥目鳥類的翅膀退化,無法用於飛行,但仍然保留了一些功能。例如,鴕鳥會使用翅膀來平衡身體,協助轉向,以及在求偶展示中發揮作用。一些鴕鳥目鳥類,如南美洲的美洲鴕,甚至可以利用翅膀在水中游泳。
在頭部和頸部特徵上,鴕鳥目鳥類也有其獨特之處。它們通常有相對較小的頭部和長而靈活的頸部。這種結構使它們能夠有效地監視周圍環境,同時也有助於覓食。鴕鳥目鳥類的喙形狀因物種而異,反映了它們不同的食性適應。
鴕鳥目鳥類的羽毛結構也與會飛的鳥類有所不同。它們的羽毛通常較為鬆軟,缺乏飛行所需的緊密結構。這種羽毛結構更適合保溫和在炎熱氣候中調節體溫。某些鴕鳥目鳥類,如鴕鳥和鴯鶓,的羽毛因其獨特的美感而在人類文化中備受青睞。
在繁殖行為上,鴕鳥目鳥類展現出多樣性。有些物種如鴕鳥採取一夫多妻制,而其他如幾維鳥則是一夫一妻制。它們通常在地面上築巢,這與它們不會飛行的特性相符。鴕鳥的蛋是所有現存鳥類中最大的,單個蛋重可達1.4公斤。代表性鴕鳥目鳥類如下:
1. 非洲大鴕鳥: 世界最大的鳥類,體高可達2.7米,重達150公斤。不能飛行,但跑步速度快。羽毛黑白相間,雄鳥黑色,雌鳥灰褐色。長頸長腿,眼睛大而明亮。腳只有兩趾。適應乾旱環境,主要分布在非洲。雜食性,以植物為主,也吃小動物。
2. 南美鴕鳥-鶆䴈 (Rhea):體型僅次於非洲鴕鳥。體高約1.5-1.7米,重達50公斤。羽毛灰褐色,頭頂和頸部較深。腳有三趾。不能飛行,但奔跑速度快。適應草原和灌木叢環境。雜食性,主要吃植物,也捕食小動物。群居性強,常成小群活動。
3. 澳洲鴯鶓:體高約1.8-2米。羽毛鬆軟,呈灰褐色。頭和頸部幾乎無羽。腳有三趾,跑步速度快。不能飛行。頭頂有角質冠。適應乾旱環境,以植物為主食,也吃昆蟲。繁殖期雄鳥孵蛋和照顧幼鳥。叫聲低沉,如打鼓聲。
4. 紐西蘭奇異鳥(kiwi):體型小,約雞大小。羽毛褐色,毛髮狀。最大特點是完全無翅,喙細長。夜行性,主要以地面昆蟲和蠕蟲為食。嗅覺極為發達。腳有四趾,爪子鋒利。因棲息地破壞和引入捕食者,現已瀕危。紐西蘭國鳥。
5. 食火雞: 巴布亞新幾內亞和澳大利亞北部特有。體型中等,似火雞大小。羽毛黑色,頭部和頸部裸露,呈藍色和紅色。腳強壯,有三趾。以腐肉和水果為食,特別喜歡吃從火山灰中挖出的烤熟的種子和昆蟲,故得名。
企鵝目
企鵝目(Sphenisciformes)是鳥類分類中一個獨特而引人注目的群體,其命名源自希臘語"sphen",意為「楔形」。這個名稱反映了企鵝身體的特殊形狀,尤其是它們流線型的體態。企鵝目的命名不僅描述了這些鳥類的外形特徵,也暗示了它們在水中游泳時的高效運動能力。最著名的有帝企鵝(體型最大)、國王企鵝、阿德利企鵝、麥哲倫企鵝。
企鵝目鳥類最顯著的特徵是它們完全適應了海洋生活。雖然它們屬於鳥類,但企鵝已經失去了飛行能力,取而代之的是出色的游泳能力。這種演化適應使得企鵝在水中如魚得水,而在陸地上則顯得笨拙。企鵝的翅膀已經演化成了類似魚鰭的結構,稱為鰭狀肢。這些鰭狀肢不僅使企鵝能夠在水中靈活地轉向和推進,還能夠在深水中保持平衡。
企鵝的身體形態是對海洋生活的完美適應。它們的身體呈流線型,有助於減少水中的阻力。厚實的脂肪層和密集的防水羽毛為它們提供了良好的保溫效果,使它們能夠在寒冷的南極和亞南極水域生存。企鵝的羽毛結構特殊,不僅防水,還能在潛水時緊貼身體,減少水的阻力。
企鵝的腿部位於身體後部,這種結構雖然使它們在陸地上行走時顯得搖搖晃晃,但在水中卻能夠提供強大的推進力。大多數企鵝物種都有黑色的背部和白色的腹部,這種配色被稱為反向隱蔽色,可以在海中為它們提供保護。從上方看,黑色的背部與深色的海水融為一體;從下方看,白色的腹部則與明亮的水面相映。
企鵝的喙部結構也適應了它們的捕食習慣。喙通常又長又尖,有些微彎曲,適合抓捕魚類和烏賊等滑溜的獵物。一些企鵝種類,如帝企鵝,還具有特殊的喙部構造,能夠過濾海水以捕獲小型甲殼類動物。
在繁殖行為上,企鵝展現出引人入勝的特點。大多數企鵝種類都是嚴格的一夫一妻制,而且通常會與同一個伴侶保持多年的關係。它們在陸地上築巢並孵化卵,這個過程中常常要面對極端惡劣的天氣條件。許多企鵝種類會形成大型繁殖群落,這不僅有利於保護後代,也形成了獨特的社會結構。
企鵝的分佈主要集中在南半球,從赤道附近的加拉帕戈斯群島一直延伸到南極大陸。不同的企鵝種類適應了不同的氣候條件,從熱帶到極地都有企鵝的身影。例如,加拉帕戈斯企鵝是唯一生活在赤道附近的企鵝種類,而帝企鵝則能夠在南極洲最寒冷的地區繁衍生息。
夜鷹目
夜鷹目(Caprimulgiformes)是鳥類分類中一個引人入勝的群體,其命名源自拉丁語"caprimulgus",字面意思是「擠山羊奶的人」。這個有趣的名稱源於一個古老的民間誤解,人們曾錯誤地認為這些鳥類會在夜間吸食山羊的奶。雖然這個說法完全是無稽之談,但這個名稱卻一直保留了下來,成為了這個鳥類目的正式學名。
夜鷹目鳥類最顯著的特徵是它們的夜行性生活方式。這些鳥類通常在黃昏和夜間活動,這種習性反映在它們的多個形態和行為特徵上。首先,夜鷹目鳥類擁有極大的眼睛,這是對夜間生活的重要適應。大眼睛能夠在微弱的光線下捕捉更多的光線,從而提高夜間視力。此外,它們的眼睛通常呈深色,這有助於減少眼內光線的散射,進一步提高夜間視力的敏銳度。
夜鷹目鳥類的喙部結構也很特別。它們的喙通常很小,但嘴裂卻異常寬大,幾乎延伸到耳朵下方。當它們張開嘴時,形成一個巨大的口腔開口,這種結構非常適合在飛行中捕捉昆蟲。在喙的周圍,許多夜鷹目鳥類還長有剛毛狀的羽毛,這些羽毛可能有助於引導飛行中的昆蟲進入口中,也可能具有觸覺功能,幫助它們在黑暗中感知周圍環境。
羽毛是夜鷹目鳥類另一個顯著的特徵。它們的羽毛通常呈現褐色、灰色或黑色的斑駁花紋,這種羽色能夠很好地融入樹皮或地面的背景中,提供出色的保護色。這種保護色不僅有助於它們在白天休息時避開天敵的注意,也能在夜間狩獵時隱藏自己的身形。
夜鷹目鳥類的飛行能力極為出色,這與它們的生活方式和捕食習慣密切相關。它們的翅膀通常又長又尖,能夠進行敏捷而無聲的飛行。這種飛行能力使它們能夠在空中靈活地追逐飛蟲,同時也能避開潛在的捕食者。一些夜鷹目鳥類,如美洲夜鷹,還以其特殊的俯衝飛行而聞名,在求偶季節,雄鳥會從高空急速下降,在接近地面時突然拉起,產生一種響亮的「嗡嗡」聲。
在腳部結構上,夜鷹目鳥類也有其獨特之處。它們的腳通常較小且脆弱,這反映了它們較少在地面活動的生活方式。有些種類的中趾有特殊的鋸齒狀結構,可能用於梳理羽毛或增強抓握能力。
夜鷹目鳥類的繁殖行為也頗具特色。大多數種類不築巢,而是直接將蛋產在地面上,依靠其羽毛的保護色來隱藏蛋和幼鳥。通常每窩只產一到兩個蛋,孵化期較長。父母鳥會輪流孵卵和照顧幼鳥,在受到威脅時,成鳥會採取各種策略來轉移潛在捕食者的注意力,如假裝受傷或發出威脅性的叫聲。
雁形目
雁形目(Anseriformes)是鳥類分類中一個重要的群體,其命名源自拉丁語"anser",意為「鵝」。這個名稱反映了鵝類在這個目中的代表性地位。雁形目包括了許多我們熟悉的水鳥,如鴨、鵝、天鵝等,這些鳥類在全球的淡水和海洋生態系統中扮演著關鍵角色。常見雁形目鳥類如下:
1. 大白鵝: 體型大,全身潔白,橙色喙和腳。頸長,姿態優雅。常見於農田、濕地和公園。性情較為溫和,但在築巢期可能具有攻擊性。
2. 鴛鴦: 雄鳥羽毛艷麗,有橙色羽扇;雌鳥褐色較樸素。常成對出現,象徵忠貞。喜歡山區溪流和湖泊,游泳和潛水能力強。
3. 綠頭鴨: 雄鳥頭部綠色有金屬光澤,頸環白色,胸部栗褐;雌鳥全身褐色帶斑點。廣泛分布於各類水域,適應性強,常見於城市公園。
4. 琵嘴鴨: 特大扁平喙部如湯匙,雄鳥頭部綠色,胸白腹栗;雌鳥褐色。常見於淺水湖泊和沼澤地,以濾食方式取食浮游生物。
5. 斑嘴鴨: 喙部黃色帶黑斑,全身羽毛灰褐色帶斑點。適應性強,廣泛分布於各類濕地環境。群居性強,常大群覓食和休息。
6. 針尾鴨: 尾部細長如針,雄鳥頭褐色,頸部白色,背灰腹白;雌鳥體色較暗淡。喜歡開闊水域,頸部修長,游泳時姿態優雅。
7. 鳳頭潛鴨: 頭頂有明顯向後的羽冠,雄鳥體色鮮艷,頭黑背白腹栗;雌鳥褐色。善於潛水捕食,常在深水區活動。
8. 小水鴨: 體型小,雄鳥頭部栗色帶綠色眼斑,胸部有黑白相間斑點。雌鳥褐色。喜歡淺水區域和沼澤地,動作靈活。
9. 鸕鶿: 體型大,全身黑色有光澤,頸部和喙細長。善於游泳和潛水捕魚。常見於沿海、湖泊等水域,曾被用於傳統漁業。
雁形目鳥類最顯著的特徵是它們對水生環境的適應。這些鳥類的身體結構明顯反映了它們的半水棲生活方式。首先,它們的腳呈蹼狀,腳趾之間有蹼膜相連,這種結構使它們能夠在水中有效地推進。腳通常位於身體後部,這種位置雖然使它們在陸地上行走時顯得笨拙,但在水中游泳和潛水時卻能提供強大的推進力。
雁形目鳥類的喙部結構也是其重要特徵。它們的喙通常又寬又扁,內側邊緣有一系列細小的角質板或齒狀突起,這種結構被稱為篩板。篩板的功能是在鳥類進食時過濾水和食物,使它們能夠有效地從水中攝取小型植物和動物。不同種類的雁形目鳥類,其喙的形狀和大小會因食性而異。例如,以植物為主食的鵝類喙部較厚實,而主要捕食魚類的秋沙鴨則有細長的鋸齒狀喙。
羽毛是雁形目鳥類另一個重要的適應特徵。它們的羽毛密集且防水,這是通過羽毛表面的油脂實現的。雁形目鳥類有特殊的腺體,稱為尾脂腺,能夠分泌油脂。鳥類會用喙將這種油脂塗抹在羽毛上,使羽毛具有防水性。這種防水羽毛不僅能保持身體乾燥,還能維持體溫,使它們能夠在寒冷的水中長時間活動。
在體型和外觀上,雁形目鳥類呈現出多樣性。它們的體型從小型的篦鴨到大型的天鵝不等。大多數雁形目鳥類的頸部相對較長,這使它們能夠在水中或淺水區覓食時更容易探及水底。許多種類還具有性二態性,即雄性和雌性在外觀上有明顯差異。例如,在許多鴨類中,雄鳥通常有鮮艷的羽毛,而雌鳥則呈現保護色。
雁形目鳥類的飛行能力也值得注意。儘管它們主要適應了水生環境,但大多數雁形目鳥類都是出色的飛行者。它們的翅膀通常又長又窄,適合長距離飛行。許多種類會進行季節性遷徙,跨越大陸和海洋。例如,加拿大雁以其長距離遷徙而聞名,每年在北美洲的繁殖地和越冬地之間往返。
在繁殖行為上,雁形目鳥類展現出多樣性。許多種類形成終身配偶關係,特別是大型的鵝和天鵝。它們通常在水邊或水中的植被中築巢,巢通常由植物材料構成,並用母鳥的絨羽襯墊。雌鳥負責孵卵,而雄鳥則守衛領地和保護家庭。一些種類,如綠頭鴨,則採取一夫多妻制。
鴴形目
鴴形目(Charadriiformes)是鳥類分類中一個多樣化且廣泛分佈的群體,其命名源自希臘語"charadrios",意為「鴴」。這個名稱反映了鴴類在這個目中的代表性地位。鴴形目包括了許多我們熟悉的濱水和海洋鳥類,如鷗、鷸、鴴、反嘴鷸等,這些鳥類在全球的海岸、濕地和內陸水域生態系統中扮演著重要角色。
鴴形目鳥類最顯著的特徵是它們對水邊環境的適應。這個目中的大多數物種都與水域有密切關係,無論是海洋、河流、湖泊還是濕地。它們的身體結構明顯反映了這種生活方式。首先,許多鴴形目鳥類有相對較長的腿,這使它們能夠在淺水區或泥灘上行走和覓食。腿的長度因物種而異,反映了它們對不同深度水域的適應。
鴴形目鳥類的喙部結構也是其重要特徵,並且在不同種類之間存在顯著差異,反映了它們多樣化的覓食策略。例如,鷸類通常有細長的喙,適合在軟泥或沙中探尋無脊椎動物;而鷗類的喙則相對較粗短,適合捕捉魚類或其他較大的獵物。一些特化的種類,如反嘴鷸,甚至進化出了上彎的喙,用於在水面上方掠食小型生物。
羽毛是鴴形目鳥類另一個重要的適應特徵。大多數種類的羽毛都具有良好的防水性,這對於在潮濕環境中生活的鳥類來說至關重要。許多鴴形目鳥類還具有季節性換羽的特徵,在繁殖季節和非繁殖季節呈現不同的羽色。例如,許多鴴和鷸在繁殖季節會換上鮮艷的羽毛,而在非繁殖季節則呈現較為樸素的羽色。
在體型和外觀上,鴴形目鳥類呈現出極大的多樣性。它們的體型從小型的鷸鴴到大型的信天翁不等。大多數鴴形目鳥類的身體結構適合飛行,有強壯的飛行肌肉和相對較長的翅膀。一些種類,如信天翁,以其出色的飛行能力而聞名,能夠在海洋上空滑翔數小時而幾乎不需拍動翅膀。
鴴形目鳥類的飛行能力通常很強。許多種類是長距離遷徙者,每年在繁殖地和越冬地之間往返數千公里。例如,北極燕鷗以其驚人的極地到極地遷徙而聞名,每年往返於北極和南極之間,創造了動物界最長的遷徙路線之一。
在繁殖行為上,鴴形目鳥類展現出多樣性。許多種類在地面上築巢,通常選擇隱蔽的位置,如草叢或沙灘上的凹陷處。一些種類,如鷗類,則會形成大型的繁殖群落。鴴形目鳥類的蛋通常具有保護色,與周圍環境相似,這有助於避免被捕食者發現。在某些種類中,如鴴類,雛鳥是早熟型,孵化後不久就能夠自行覓食。
鴴形目鳥類在生態系統中扮演著多重角色。作為濱水和海洋生態系統的重要成員,它們對維持這些環境的生態平衡起著關鍵作用。許多種類是重要的捕食者,控制著小型魚類和無脊椎動物的數量。同時,它們也是許多捕食者的獵物,構成了食物鏈的重要環節。此外,一些鴴形目鳥類,如鷗類,還通過其糞便為海洋島嶼提供重要的營養輸入。
鷸科 (Scolopacidae)
鷸:主要屬於鷸科(Scolopacidae),這些鳥通常體型中等到較大,擁有修長的腿和細長的嘴,適合在各種濕地環境中覓食。鷸的嘴往往較長,有些種類的嘴還略微彎曲,這使它們能夠深入泥中探測食物。它們的羽毛通常呈現褐色系,具有良好的保護色,能夠很好地融入環境。
鷸類在覓食時常可見到特徵性的點頭或身體上下擺動的動作,這有助於它們定位獵物。飛行時,鷸類通常保持頸部伸直的姿態。它們的叫聲多樣,有的低沉,有的尖銳,為濕地增添了豐富的音景。
許多鷸類是長距離遷徙鳥,它們能夠進行令人驚嘆的洲際飛行。它們偏好棲息在濕地、泥灘和沼澤地等水邊環境,在這些地方,你常能看到它們成群結隊地在淺水中覓食的景象。鷸類在濕地生態系統中扮演著重要角色,是環境變化的敏感指標。
1. 青足鷸: 中小型涉禽,體長約22-25厘米。上體灰褐色,下體白色,繁殖期胸部有褐色斑點。腿部黃綠色,喙細長微向上彎。常見於泥灘和淺水區域。警戒性強,受驚時發出尖銳的"tluit"叫聲。
2. 鷹斑鷸: 中型涉禽,體長約27-30厘米。上體褐色帶有白色斑點,下體白色有褐色斑紋。喙粗壯微下彎,腿部黃綠色。常單獨或小群活動於泥灘和濕草地。飛行時翅膀有明顯白色條帶。
3. 黑腹濱鷸: 小型涉禽,體長約18-20厘米。繁殖羽上體灰褐色,下體黑色;非繁殖羽全身灰色。喙短而直,腿黑色。常成群出現在泥灘和淺水區。飛行快速,常呈緊密編隊。
4. 丘鷸: 中小型涉禽,體長約25-27厘米。上體褐色帶有複雜斑紋,下體白色。頭頂有明顯的黑色縱紋。喙長而直,腿部黃綠色。常見於草地和農田。飛行時發出特徵性的"嘟-嘟-嘟"叫聲。
5. 長趾濱鷸: 小型涉禽,體長約15-18厘米。腳趾特別長,適應在浮水植物上行走。上體褐色帶有斑點,下體白色。常見於沼澤和蘆葦叢中。行走時動作輕盈,看似在水面上漂浮。
6. 尖尾濱鷸: 小型涉禽,體長約20-22厘米。尾部中央羽毛較長,呈尖形。上體灰褐色,下體白色。繁殖期胸部有褐色斑點。常見於泥灘和濕地邊緣。群居性強,受驚時整群快速起飛。
7. 大杓鷸: 大型涉禽,體長約50-60厘米。最顯著特徵是極長的向下彎曲的喙。全身褐色帶有斑點,腿部灰藍色。常見於泥灘和濕地。用長喙在泥中探尋螃蟹和蠕蟲。叫聲低沉悅耳。
8. 中杓鷸: 中型涉禽,體長約40-45厘米。與大杓鷸相似但體型較小,喙彎曲程度較小。全身褐色帶有斑點,腿部藍灰色。常見於海岸和內陸濕地。群居性強,遷徙時常成群出現。
9. 磯鷸: 小型涉禽,體長約20-22厘米。上體褐色帶有暗斑,下體白色帶有褐色斑點。尾部常上下擺動,為其特徵之一。喜歡在岩石海岸和溪流邊活動。動作敏捷,善於在岩石間穿梭。
鴴科 (Charadriidae)
相較於鷸,鴴通常體型較小,腿相對短一些。它們的嘴短而直,非常適合在地面或淺水區域撿拾食物。鴴類的羽毛常常呈現出醒目的pattern,許多種類有著標誌性的黑白相間頸環或胸帶,這些特徵在繁殖期尤為明顯。
鴴類的行為特徵之一是其獨特的停頓-跑動方式,這種動作有助於它們發現和捕捉獵物。飛行時,鴴類的頸部略微縮回,呈現出與鷸不同的飛行剪影。它們的叫聲通常清脆悅耳,在開闊的環境中傳播很遠。
雖然許多鴴類也進行季節性遷徙,但相比鷸類,它們中有更多種類傾向於在某一地區定居。鴴類的棲息地選擇較為多樣,除了濕地外,也常見於乾燥的開闊地帶,如草原、沙地甚至農田。這種適應性使得鴴類在不同的生態系統中都扮演著重要角色。
鴴類的多樣性和適應性使它們成為鳥類觀察者的熱門目標,也是生態研究中重要的研究對象。它們優雅的身姿和有趣的行為為自然世界增添了無窮的魅力。常見的鴴科鳥類如下:
1. 金斑鴴: 中型鴴類,體長約27-30厘米。繁殖羽上體金褐色帶黑斑,下體黑色;非繁殖羽較淡。頭部、頸部和胸部有明顯的白色斑塊。喙短黑色,腿較長。常見於開闊草地、荒地和農田。飛行時腋下黑色,翅膀有白色條紋。叫聲清脆悅耳。
2. 鳳頭麥雞: 中型鴴類,體長約28-31厘米。最顯著特徵是頭頂有黑色羽冠。上體褐色,下體白色,頸部有黑色環帶。喙短黑色,腿黃色。常見於開闊草地、農田和濕地邊緣。行為活潑,受驚時發出尖銳的"pee-wit"叫聲。繁殖期會做出誇張的空中求偶表演。
3. 小辮鴴: 小型鴴類,體長約16-18厘米。頭頂有短小黑色羽辮,黑色貫眼紋明顯。上體褐色,下體白色,頸部有不完整的黑色環帶。喙短黑色,腿黃色。常見於沙灘、泥灘和濕地邊緣。動作靈活,跑動時斷續前進。築巢於地面,常做出受傷鳥狀態以引開天敵。
4. 環頸鴴: 小型鴴類,體長約17-19厘米。頸部有明顯的黑色環帶,上體褐色,下體白色。額頭白色,眼後有黑色斑塊。喙短黑色,基部橙黃色,腿橙黃色。廣泛分布於各類濕地環境。警惕性高,受驚時快速奔跑或低空飛行。常做出假受傷動作以保護巢穴。
5. 劍鴴: 中型鴴類,體長約25-28厘米。上體灰褐色,下體白色,胸部有一道黑色橫帶。頭部和頸部有複雜的黑白相間花紋。喙長而粗壯,呈黑色。腿較長,呈黃綠色。常見於泥灘、沙灘和鹽沼。覓食時快速奔跑,突然停下啄食。群居性強,常成小群活動。
6. 紅胸鴴: 小型鴴類,體長約14-16厘米。繁殖羽胸部為鮮艷的磚紅色,頭部和頸部有黑白相間花紋;非繁殖羽較淡。上體褐色,下體白色。喙短黑色,腿黑色。多見於海岸和河口濕地。動作敏捷,跑動時如同小風車。遷徙時可進行長距離飛行。
7. 蒙古沙鴴: 小型鴴類,體長約18-21厘米。繁殖羽頭部和胸部有醒目的栗色斑塊,眉斑和喉部白色;非繁殖羽較淡。上體褐色,下體白色。喙短黑色,腿深色。常見於沙質海岸和內陸沙地。適應乾旱環境,能在高溫沙地上奔跑覓食。巢常建在植物遮蔽下。
8. 鐵嘴沙鴴: 中型鴴類,體長約23-26厘米。全身灰褐色,腹部較淡。最顯著特徵是粗壯的紅色喙。眼圈和腿部也呈紅色。常見於岩石海岸和礁石區。善於在礁石間穿梭覓食,以貝類和甲殼類為食。叫聲洪亮,類似口哨聲。適應性強,也見於內陸地區。
9. 長嘴鴴: 中型鴴類,體長約23-25厘米。最顯著特徵是細長的黑色喙。上體褐色,下體白色,胸部有一道黑色橫帶。頭部和頸部有複雜的黑白相間花紋。腿較長,呈黑色。常見於泥灘和淺水區域。用長喙在泥中探尋食物,如蠕蟲和甲殼類。警惕性高,受驚時發出尖銳的叫聲。
10. 灰頭麥雞: 中型鴴類,體長約28-31厘米。頭部和頸部為淺灰色,與褐色的身體形成對比。胸部有黑色橫帶,腹部白色。喙短黑色,腿黃色。常見於開闊草地、農田和濕地邊緣。行為類似鳳頭麥雞,但叫聲較低沉。繁殖期會進行空中求偶表演,發出特殊的嗡嗡聲。
鷗科 (Laridae)
鷗科(Laridae)包括鷗和燕鷗兩大類群,它們是海洋和沿海環境中最常見的鳥類之一,也常見於內陸的大型水體周圍。 鷗科鳥類通常體型中等到較大,擁有強健的身體和長而尖的翅膀,使它們能夠在空中靈活飛行並能夠滑翔長距離。它們的嘴通常又長又強壯,略微彎曲,適合捕捉魚類和其他水生生物。腳上有蹼,有助於在水面游泳和起飛。 在羽毛方面,成年鷗類通常以白色、灰色和黑色為主。許多種類在非繁殖期和繁殖期有不同的羽色。年輕的鷗往往呈現褐色或灰色的斑駁pattern,需要幾年時間才能發育出成鳥的羽毛。 鷗科鳥類以其多樣的叫聲而聞名,從悅耳的鳴叫到刺耳的尖叫都有。這些聲音在海濱和港口形成了標誌性的聲景。它們通常群居性強,常在覓食地和繁殖地形成大型群落。 在生態習性方面,鷗類表現出驚人的適應能力和智慧。它們是機會主義覓食者,食物範圍廣泛,包括魚類、軟體動物、甲殼類,甚至人類丟棄的食物。一些種類還會跟隨漁船或在垃圾填埋場覓食,顯示出對人類活動的適應。 繁殖時,鷗類通常選擇在沿海岩礁、沙灘或島嶼上築巢。它們的巢通常很簡單,只是地上的一個淺窩。許多種類具有強烈的領地意識,會積極保護自己的巢區。常見的鷗科鳥類如下:
1. 銀鷗: 大型鷗類,體長約55-65厘米。成鳥背部和翅膀淺灰色,其餘部分白色。翅尖黑色帶白點。喙黃色,腿粉紅色。冬季頭部有灰色斑紋。常見於海岸、港口和內陸水域。適應性強,也出現在城市地區。叫聲響亮,常發出"k-yo-oh"的聲音。
2. 紅嘴鷗: 中型鷗類,體長約40-45厘米。成鳥全身白色,背部和翅膀淺灰色。最顯著特徵是鮮紅色的喙和腳。眼圈紅色。冬季頭部可見灰色斑點。常見於內陸湖泊、河流和海岸。飛行優雅,常低空盤旋。繁殖期頭部會變為黑色。群居性強,常大群聚集。
3. 黑尾鷗: 中型鷗類,體長約40-45厘米。成鳥背部和翅膀深灰色,頭部、頸部和腹部白色。尾部有明顯的黑色橫帶。喙紅色帶黑色尖端,腿深紅色。冬季頭部有灰色斑紋。常見於海岸和河口地區。飛行靈活,善於俯衝捕魚。繁殖期頭部變為黑色。
4. 海鷗: 大型鷗類,體長約60-70厘米。成鳥背部和翅膀暗灰色,其餘部分白色。翅尖黑色。喙黃色帶紅色斑點,腿粉紅色。冬季頭部有褐色斑紋。廣泛分布於海岸、港口和內陸水域。適應性極強,常在人類活動區覓食。叫聲低沉有力。
5. 燕鷗: 中小型鷗科鳥類,體長約33-38厘米。成鳥上體淺灰色,下體白色。頭頂黑色,尾部分叉。喙細長,呈紅色帶黑色尖端。腿短小,紅色。飛行敏捷,常見低空掠水捕魚。繁殖期頭頂變為全黑色。群居性強,常在沙洲或礁石上築巢。
6. 三趾鷗: 小型鷗類,體長約35-40厘米。成鳥上體淺灰色,下體白色。翅膀邊緣有獨特的黑白相間花紋。腿短,只有三趾,呈黑色。喙黃色。常見於開闊海域,很少在陸地上停留。飛行能力極強,能在風浪中穿梭自如。繁殖期頭部變為淺灰色。
7. 鳳頭燕鷗: 中型燕鷗,體長約40-45厘米。成鳥上體淺灰色,下體白色。頭頂黑色並延伸成羽冠。喙粗壯,鮮黃色。腿黑色。尾部分叉不明顯。常見於海岸、河口和內陸水域。飛行穩定有力,捕魚時從高處俯衝入水。繁殖期嘴變為橙紅色。群居性強。
鸚 形目
鸚形目(Psittaciformes)是鳥類分類中一個引人注目且廣受喜愛的群體,其命名源自古希臘語"psittakos",意為「鸚鵡」。這個名稱直接反映了鸚鵡在這個目中的核心地位。鸚形目包括了我們熟知的鸚鵡、鸚哥、摩卡鸚鵡等鳥類,它們以鮮艷的羽毛、彎曲的喙和高度智慧而聞名於世。
常見的鸚形目鳥類包括了多個廣為人知的種類。其中最為人熟知的可能是灰鸚鵡,它以出色的模仿人類語言能力而聞名。金剛鸚鵡是體型最大的鸚鵡之一,羽毛鮮艷,常見於熱帶雨林。虎皮鸚鵡體型較小,性格活潑,是受歡迎的寵物鳥。澳洲的鳳頭鸚鵡以其獨特的冠羽而聞名。此外,還有常見於南美洲的金剛鸚鵡、非洲的長尾鸚鵡,以及分佈廣泛的牡丹鸚鵡等。
鸚形目鳥類最顯著的特徵是它們獨特的喙部結構。這些鳥類擁有強壯、彎曲的喙,上喙能夠靈活地與下喙活動。這種喙部結構不僅使它們能夠輕易地破開堅果和種子,還能夠用作「第三隻腳」,幫助它們在樹枝間攀爬。鸚鵡的舌頭也很特別,肉質且靈活,有助於操縱食物和發出複雜的聲音。
另一個顯著特徵是鸚形目鳥類的腳部結構。它們的腳呈現對趾型,即兩趾向前,兩趾向後。這種腳部結構使它們能夠牢固地抓握樹枝,並在樹上靈活地移動。許多鸚鵡物種還能用腳將食物送到嘴邊,展現出令人驚嘆的靈巧性。
鸚形目鳥類以其鮮艷的羽毛而聞名。它們的羽色範圍從鮮豔的紅色、藍色、綠色到較為樸素的灰色不等。這些色彩不僅美麗,還在鸚鵡的社交行為和求偶過程中扮演重要角色。一些物種,如金剛鸚鵡,甚至能夠控制面部的羽毛,用來表達情緒。
智能是鸚形目鳥類另一個顯著特徵。這些鳥類以其高度發達的認知能力而聞名,能夠解決複雜問題、使用工具,甚至學習人類語言。例如,灰鸚鵡亞歷克斯(Alex)曾展示出理解數字概念和簡單語言的能力,震驚了科學界。這種智能與鸚鵡相對較大的大腦有關,特別是負責高級認知功能的前腦區域。
在社交行為方面,鸚形目鳥類通常高度群居。它們常常形成大型群體,在覓食和休息時保持緊密聯繫。這種社交性也反映在它們的交流方式上,鸚鵡能夠發出各種複雜的叫聲和鳴叫,用於在群體中傳遞信息。
鸚形目鳥類的繁殖行為也很有特色。大多數物種形成終身配偶關係,展現出高度的配偶忠誠度。它們通常在樹洞或岩洞中築巢,有些物種甚至會使用工具來改造巢穴。鸚鵡的繁殖週期較長,雛鳥需要較長時間的親代照顧才能獨立。
隼 形目
隼形目(Falconiformes)是鳥類分類中一個引人注目的群體,其命名源自拉丁語"falco",意為「隼」。這個名稱直接反映了隼類在這個目中的核心地位。隼形目包括了許多著名的猛禽,如隼、鷹、鷹隼等,這些鳥類以其銳利的視力、敏捷的飛行能力和捕獵技巧而聞名。
15種常見的隼形目鳥類包括:遊隼(Peregrine Falcon)、紅隼(Common Kestrel)、燕隼(Hobby)、灰背隼(Merlin)、矛隼(Lanner Falcon)、獵隼(Saker Falcon)、白隼(Gyrfalcon)、紅腳隼(Red-footed Falcon)、美洲隼(American Kestrel)、禿鷹(Bald Eagle)、金鵰(Golden Eagle)、蒼鷹(Goshawk)、鵟(Common Buzzard)、鳶(Black Kite)和魚鷹(Osprey)。這些鳥類在全球各地廣泛分佈,每種都有其獨特的特徵和生態位。
隼形目鳥類最顯著的特徵是它們的喙部和爪子。它們擁有強壯、彎曲的喙,上喙末端呈鉤狀,非常適合撕裂獵物的肉。這種喙部結構使它們能夠有效地處理各種獵物,從小型哺乳動物到鳥類和昆蟲。隼形目鳥類的爪子同樣強壯而尖銳,通常有四趾,其中三趾向前,一趾向後。這種爪子結構使它們能夠牢固地抓住獵物和棲息地。
另一個顯著特徵是隼形目鳥類的視力。它們擁有極其敏銳的視力,這在捕獵時起著關鍵作用。例如,遊隼能夠從高空準確地識別並鎖定地面上的獵物。這種出色的視力與它們相對較大的眼睛和高度發達的視覺皮層有關。
隼形目鳥類的飛行能力也令人驚嘆。它們的翅膀通常又長又窄,適合高速飛行和靈活操控。不同物種的翅膀形狀略有不同,反映了它們的捕獵策略。例如,遊隼的翅膀呈尖銳的三角形,適合高速俯衝;而鷹的翅膀則較寬,適合滑翔和盤旋。
在體型和外觀上,隼形目鳥類呈現出多樣性。它們的體型從小型的紅隼到大型的禿鷹不等。羽色通常較為樸素,以褐色、灰色和白色為主,這有助於它們在捕獵時保持隱蔽。然而,也有一些物種,如禿鷹,具有醒目的頭部羽色。
隼形目鳥類的捕獵行為是它們最引人注目的特徵之一。不同物種有不同的捕獵策略。例如,遊隼以其高速俯衝聞名,能夠達到每小時320公里的速度,是地球上最快的動物之一。而鷹則通常採用盤旋和突然俯衝的策略。魚鷹則特化於捕捉魚類,能夠直接俯衝入水捕獵。
在繁殖行為上,隼形目鳥類通常形成穩定的配偶關係。它們常在高處築巢,如懸崖、高樹或建築物上。巢通常由樹枝構成,並用柔軟的材料襯墊。大多數物種每年只繁殖一次,產卵數量較少,但會投入大量精力照顧幼鳥。
隼形目鳥類在生態系統中扮演著頂級捕食者的角色。它們通過控制獵物數量來維持生態平衡。例如,隼類對控制鼠類和小型鳥類的數量起著重要作用,而鷹則可能捕食較大型的哺乳動物。此外,一些物種如禿鷹,還通過清理動物屍體來發揮生態淨化作用。
在人類文化中,隼形目鳥類佔有重要地位。它們常被視為力量、自由和勇氣的象徵。在許多文化中,這些鳥類與神話和傳說有關。例如,禿鷹是美國的國鳥,象徵著力量和自由。在一些地區,隼類還被用於傳統的獵鷹活動。
隼形目的分類學研究一直在不斷深入。隨著分子生物學技術的發展,科學家們對這個群體的演化歷史和物種關係有了新的認識。例如,近期的研究表明,鷹隼可能與隼類的親緣關係比以前認為的更近。這些發現不斷更新我們對鳥類演化的理解。
隼形目鳥類為我們提供了研究適應性進化、捕食行為和生態學的絕佳範例。它們展示了鳥類如何發展出高度專門化的捕獵技能和身體結構。這種獨特的生態位使得隼形目鳥類成為生態學、行為學和保護生物學研究的重要對象。通過對這些迷人鳥類的深入研究,我們不僅能更好地理解頂級捕食者在生態系統中的作用,也能為瀕危物種的保護提供重要的科學依據。
鴷 形目
鴷形目(Piciformes)是鳥類分類中一個獨特而多樣的群體,其命名源自拉丁語"picus",意為「啄木鳥」。這個名稱反映了啄木鳥在這個目中的核心地位。鴷形目包括了許多引人注目的鳥類,如啄木鳥、巨嘴鳥和鬚鴷等,這些鳥類以其獨特的形態、行為和生態位而聞名。
東亞常見的鴷形目(Piciformes)鳥類及其特徵:
- 大赤啄木鳥 (Dendrocopos major): 體長約23cm,黑白相間,雄鳥後腦有紅斑。喜歡在樹幹上啄食昆蟲,叩擊聲響亮。
- 小啄木鳥 (Dendrocopos minor): 體長約15cm,是最小的啄木鳥之一。黑白相間,雄鳥頭頂紅色。動作敏捷,常在細枝間活動。
- 綠啄木鳥 (Picus canus): 體長約30cm,全身綠色,頭部灰色,雄鳥前額紅色。常在地面覓食,喜歡吃蟻。
- 大斑啄木鳥 (Dendrocopos major): 體長約23cm,黑白相間,腹部淡紅色,雄鳥後腦有紅斑。適應性強,分佈廣泛。
- 五色鳥 (Megalaima nuchalis): 體長約20cm,羽毛色彩豐富,包括紅、藍、綠、黃、黑等。主要吃果實和昆蟲。
這些鳥類在生態系統中扮演著重要角色,特別是在控制森林害蟲方面。
鴷形目鳥類最顯著的特徵是它們的喙部結構。這些鳥類通常擁有強壯、尖銳的喙,特別適合啄木、鑿洞和捕捉昆蟲。啄木鳥的喙更是進化出了特殊的減震結構,使它們能夠長時間啄擊樹木而不受傷。巨嘴鳥則以其巨大而輕盈的喙而聞名,這種喙結構有助於它們在樹冠層中平衡身體並採食果實。
另一個顯著特徵是鴷形目鳥類的腳部結構。大多數物種擁有攀爬型腳,通常是兩趾向前、兩趾向後的排列(攀禽足),這種結構使它們能夠牢固地抓住樹幹和樹枝,在垂直表面上自如移動。啄木鳥的爪子特別發達,能夠緊緊抓住樹皮。
鴷形目鳥類的舌頭結構也非常獨特。啄木鳥的舌頭極長,能夠伸出喙外很長的距離,用來探索樹洞並捕捉昆蟲。這種舌頭甚至能夠纏繞在頭骨周圍,當不使用時可以收縮回去。巨嘴鳥的舌頭則相對較短,但具有刷子狀的結構,有助於處理果實。
在外觀上,鴷形目鳥類呈現出豐富的多樣性。啄木鳥通常有黑白相間的羽毛,常常帶有紅色或黃色的裝飾性羽毛。巨嘴鳥則以其鮮艷的羽色和巨大的喙而聞名。鬚鴷通常體型較小,但羽色豐富多彩。
鴷形目鳥類的行為特徵也很有趣。啄木鳥以其特殊的啄木行為而聞名,不僅用於覓食,也用於築巢和交流。它們通過有節奏的敲擊來標記領地和吸引配偶。巨嘴鳥則以其獨特的鳴叫聲和社交行為而著稱,常常成群結隊活動。
在繁殖行為上,鴷形目鳥類通常在樹洞中築巢。許多物種,特別是啄木鳥,會自己鑿洞築巢。這些巢穴不僅用於繁殖,還為森林生態系統中的其他動物提供了重要的棲息場所。大多數鴷形目鳥類都是一夫一妻制,雙親共同孵卵和餵養幼鳥。
鴷形目鳥類在生態系統中扮演著重要角色。啄木鳥通過控制樹皮下昆蟲的數量來維護森林健康。它們創造的樹洞為許多其他動物提供了棲息地。巨嘴鳥和鬚鴷則是重要的種子傳播者,對維持熱帶森林的生物多樣性起著關鍵作用。
鴞 形目
鴞形目(Strigiformes)是鳥類分類中一個引人入勝的群體,其命名源自拉丁語"strix",意為「貓頭鷹」。這個名稱直接反映了貓頭鷹在這個目中的核心地位。鴞形目包括了我們熟知的貓頭鷹和鴟鴞,這些鳥類以其獨特的外貌、夜行習性和高度專門化的捕獵技能而聞名。中文"鴞"是一個象形字,描繪了貓頭鷹的形象。這個字的上半部分"留"像是貓頭鷹的大眼睛。
- 領角鴞 (Otus lettia): 體長約23cm,耳狀羽簇明顯。羽色多變,從灰褐到紅褐不等。眼睛黃色。夜行性,發出低沉的"咕-咕"叫聲。常見於森林和城市公園。
- 黃嘴角鴞 (Otus spilocephalus): 體長約19cm,體型小巧。羽毛褐色帶斑點,嘴喙黃色。耳羽不明顯。喜歡棲息在山地森林中。叫聲像貓叫,故又稱"貓頭鷹"。
- 褐林鴞 (Strix leptogrammica): 體長約45cm,無耳羽。全身褐色帶有細密的橫紋。眼睛深褐色。棲息於茂密的森林中,以小型哺乳動物為食。
- 鵂鶹 (Ninox scutulata): 體長約30cm,體型修長。上體褐色,下體有縱紋。眼睛黃色。晝夜皆活動,常發出"咕-咕"的叫聲。廣泛分佈於各類林地。
- 長耳鴞 (Asio otus): 體長約36cm,耳羽長而明顯。羽毛褐色帶有黑色縱紋。眼睛橙黃色。夜行性,白天隱藏在茂密的樹枝間。
- 短耳鴞 (Asio flammeus): 體長約38cm,耳羽短而不明顯。羽毛淡褐色帶有暗色斑紋。眼睛黃色,周圍有明顯的黑色眼圈。常在開闊地帶活動。
- 倉鴞 (Tyto alba): 體長約34cm,面盤呈心形。上體金黃色帶灰色斑點,下體白色。眼睛黑色。常棲息於建築物中,故又稱"教堂貓頭鷹"。
- 斑頭鴞 (Glaucidium cuculoides): 體長約23cm,體型小巧。頭部有明顯的白色斑點。背部褐色,腹部有縱紋。晝夜皆活動,常發出清脆的叫聲。
- 貓頭鷹 (Bubo bubo): 體長約70cm,是最大的鴞形目鳥類之一。耳羽明顯,全身褐色帶有黑色斑紋。眼睛橙紅色。主要捕食中型哺乳動物和鳥類。
- 鷹鴞 (Ketupa flavipes): 體長約60cm,體型大而強壯。羽毛褐色帶有黑色縱紋,腳部無羽毛。眼睛黃色。常在水邊活動,以魚類和兩棲類為食。
鴞形目鳥類最顯著的特徵是它們的頭部結構。這些鳥類通常擁有大而圓的頭部,配有一張扁平的「臉盤」,這種結構有助於收集聲音並引導到耳朵。它們的眼睛大而前置,提供出色的立體視覺,特別適合在低光條件下捕獵。許多種類還具有明顯的耳羽,雖然這些「耳朵」實際上只是裝飾性的羽毛。
另一個顯著特徵是鴞形目鳥類的聽覺能力。它們的耳朵通常不對稱排列,一個稍高一個稍低,這種構造使它們能夠精確定位聲音的來源。結合其特殊的頭部羽毛結構,貓頭鷹能夠在完全黑暗的環境中僅憑聲音就能捕獲獵物。
鴞形目鳥類的羽毛結構也很特別。它們的飛行羽毛邊緣有鋸齒狀結構,這使得它們能夠無聲飛行,是夜間捕獵的重要適應。羽毛通常呈現保護色,如褐色、灰色或白色,有助於它們在白天休息時隱藏自己。
在行為方面,鴞形目鳥類以其夜行性而著稱。大多數種類在黃昏和夜間活動,利用其敏銳的感官捕獵小型哺乳動物、鳥類和昆蟲。然而,也有一些種類,如短耳鴞和鷹鴞,會在白天活動。
鴞形目鳥類的捕獵技巧高度專門化。它們通常從高處觀察,然後以迅速而無聲的飛行撲向獵物。強大的爪子和彎曲的喙是它們有效捕獵和處理獵物的工具。一些種類,如倉鴞,能夠僅憑聲音就能在完全黑暗中精確定位並捕捉老鼠。
在繁殖行為上,鴞形目鳥類通常形成穩定的配偶關係,有些甚至終身配對。它們常常利用樹洞、岩洞或廢棄的建築物築巢。有趣的是,很多種類不會自己建造巢,而是利用其他鳥類遺棄的巢或天然的凹洞。雌鳥負責孵卵,而雄鳥則提供食物。
鴞形目鳥類在生態系統中扮演著重要的角色。作為頂級捕食者,它們有助於控制小型哺乳動物和鳥類的數量,從而維持生態平衡。同時,它們的存在也是生態系統健康的重要指標。
雞形目
雞形目(Galliformes)是一個重要的目,包含了許多常見的地棲鳥類。雞形目的命名源自拉丁語"gallus",意為"公雞"。這個名稱反映了該目中許多鳥類與家雞的親緣關係及相似特徵。雞形目鳥類通常具有強壯的喙和爪,適合在地面覓食和刨土。
常見的雞形目鳥類及其主要特徵:
- 家雞(Gallus gallus domesticus): 最廣泛飼養的家禽,體型多樣,羽毛豐富多彩。
- 野雞(Gallus gallus): 家雞的野生祖先,棲息於東南亞叢林,雄鳥具有鮮艷的羽毛和長尾。
- 孔雀(Pavo cristatus): 雄鳥以華麗的尾屏著稱,常見於印度次大陸。
- 火雞(Meleagris gallopavo): 北美原產,頭頸裸露,雄鳥有特徵性的胸前毛髯。
- 山雞(Chrysolophus pictus): 原產中國,雄鳥羽毛鮮艷,有金黃色冠羽。
- 鵪鶉(Coturnix japonica): 體型小巧,善於奔跑,常被飼養作食用或寵物。
- 松雞(Tetrao urogallus): 歐亞大陸針葉林中的大型鳥類,雄鳥黑色,有特殊的求偶展示。
- 雉雞(Phasianus colchicus): 原產亞洲,已被引入世界多地,雄鳥有鮮艷的羽毛和長尾。
- 鷓鴣(Bambusicola): 亞洲南部和東南部的林地鳥類,體型較小,善於隱蔽。
- 竹雞(Bambusicola thoracicus): 中國南部常見,喜歡在竹林和灌木叢中活動。
這些雞形目鳥類雖然在外形和生活習性上有所差異,但都具有一些共同特徵。它們大多具有強壯的腿和爪,適合在地面行走和刨土覓食。喙部通常短而堅硬,適合啄食種子、漿果和小型動物。許多雞形目鳥類存在明顯的性二型現象,雄性通常比雌性體型更大,羽毛更鮮艷。這些鳥類多數營巢於地面,幼鳥孵化後不久即能跟隨父母活動,屬於早熟型。
雞形目鳥類在生態系統中扮演重要角色,既是許多捕食者的獵物,也通過食性影響植被分佈。同時,它們與人類關係密切,不僅是重要的食物來源,還在文化、藝術和宗教中佔有重要地位。隨著環境變化和人類活動的影響,一些雞形目鳥類面臨棲息地喪失和過度捕獵的威脅,需要加強保護措施以確保其種群的可持續發展。
人類對雞形目鳥類的研究不僅限於分類學,還涉及行為學、生態學、進化生物學等多個領域。這些研究不僅有助於我們更好地瞭解和保護這些鳥類,也為農業生產、疾病防控和生物多樣性保護提供了重要的科學依據。
鶴形目
在眾多鳥類目中,鶴形目(Gruiformes)是一個引人注目的群體。鶴形目的命名源自拉丁文"grus",意為"鶴"。這個名稱反映了該目中最具代表性的鳥類—鶴科鳥類的特徵。鶴形目鳥類多為中大型水鳥或濕地鳥類,具有長腿、長頸和尖喙等共同特徵,適應了在沼澤、草原和淺水區域生活的需求。
常見的鶴形目鳥類及其主要特徵:
- 丹頂鶴(Grus japonensis): 體型優雅,羽毛主要為白色,頭頂有鮮明的紅色冠羽,被視為長壽和忠誠的象徵,在東亞文化中具有重要地位。
- 灰鶴(Grus grus): 歐亞大陸廣泛分佈的鶴類,體色以灰色為主,頭部有黑白相間的圖案,飛行時頸部伸直。
- 沙丘鶴(Grus canadensis): 北美洲常見鶴類,體型較小,灰色羽毛,額頭有紅色斑塊,善於適應各種棲息地。
- 白枕鶴(Grus vipio): 東亞鶴類,頸部白色,頭頂紅色,背部灰色,喜歡在濕地和農田覓食。
- 彩鷸(Rostratula benghalensis): 體型小巧,羽毛華麗,雌鳥比雄鳥更鮮艷,常見於亞洲和非洲的濕地。
- 秧雞(Rallus aquaticus): 體型細長,善於在茂密的水生植物中穿梭,羽毛為褐色和灰色,有助於偽裝。
- 水雞(Gallinula chloropus): 全身黑色,額頭有紅色盾片,腳趾細長,適合在水面植物上行走。
- 白腰草鷸(Tringa ochropus): 屬於鷸科,但被歸類於鶴形目,體型中等,上體暗褐色,飛行時可見明顯的白色尾楔。
- 白鶴(Grus leucogeranus): 全身潔白,只有飛羽呈黑色,面部裸露部分為紅色,是極度瀕危物種。
- 蒼鷺(Ardea cinerea): 雖然傳統上被歸類為鷺科,但在某些分類系統中被納入鶴形目,體型大,羽毛以灰色為主。
- 董雞(Gallicrex cinerea): 體型較大的秧雞科鳥類,雄鳥繁殖期時額頭有紅色肉垂,常見於稻田和沼澤地。
這些鶴形目鳥類雖然在外形和生活習性上各有特點,但都展現出一些共同的特徵。它們大多具有修長的腿和頸,適應了在水邊或濕地環境中覓食和生活的需求。許多種類都有發達的趾,能夠在軟泥和水生植物上自如行走。鶴形目鳥類的喙形狀多樣,反映了它們不同的食性適應。有些種類如鶴科鳥類,喙較長而直,適合在淺水中捕捉魚類和兩棲動物;而秧雞科鳥類的喙則相對短小,更適合啄食植物種子和小型無脊椎動物。
隼 形目
隼形目(Falconiformes)是鳥類分類中一個引人注目的群體,其命名源自拉丁文"falco",意為"隼"。這個名稱反映了該目中最具代表性的鳥類—隼科鳥類的特徵。隼形目鳥類以其敏捷的飛行能力、銳利的視力和捕獵技巧而聞名,是空中的頂級掠食者。
常見的隼形目鳥類及其主要特徵:
- 遊隼(Falco peregrinus)是隼形目中最著名的成員之一,以其高速俯衝捕獵而聞名。這種中型猛禽全球分佈廣泛,羽毛呈灰藍色,頭部有明顯的鬍鬚斑。遊隼的飛行速度可達每小時320公里,是地球上速度最快的動物之一。
- 紅隼(Falco tinnunculus)是一種小型隼,常見於開闊地區和城市環境。雄鳥頭部和尾部呈藍灰色,背部紅褐色帶黑斑,雌鳥則全身褐色帶斑點。紅隼以其特殊的懸停飛行方式捕捉地面的小型獵物而聞名。
- 燕隼(Falco subbuteo)是一種優雅的中小型隼,以捕食飛行中的昆蟲和小型鳥類而著稱。它們有細長的翅膀和尾巴,飛行敏捷,常在黃昏時分活動。
- 草原隼(Falco naumanni)是一種小型隼,主要棲息在乾旱和半乾旱地區。雄鳥頭部和尾部呈灰藍色,背部紅褐色無斑點,與紅隼相似但體型更小。
- 白隼(Falco rusticolus)是最大的隼科鳥類,主要分佈在北極地區。羽毛以白色為主,混有灰色或褐色斑紋,適應了極地環境。
- 日本松雀鷹(Accipiter gularis)雖然屬於鷹科,但在某些分類系統中被歸入隼形目。這種小型猛禽主要分佈在東亞,以捕食小型鳥類為主。
- 鳳頭蒼鷹(Accipiter trivirgatus)是一種中型林鷹,主要分佈在南亞和東南亞的森林地區。它有特徵性的冠羽和強壯的爪,善於在茂密的林間捕獵。
- 蜂鷹(Pernis ptilorhynchus)是一種特化的猛禽,主要以蜂類和黃蜂幼蟲為食。它們有特殊的羽毛結構保護眼睛免受蜂刺傷害。
- 黑鳶(Milvus migrans)是一種適應性極強的中型猛禽,常見於人類聚居地附近。它們以其叉形尾巴和滑翔飛行而易於識別。
- 赤腹鷹(Accipiter soloensis)是一種小型猛禽,主要分佈在東亞。繁殖期間,雄鳥腹部呈現明顯的紅褐色,是其顯著特徵。
- 鵟(Buteo buteo)是一種中型猛禽,羽色變化多樣,從淺褐色到深褐色不等。它們常見於開闊地區,以滑翔飛行尋找獵物。
- 禿鷹(Aegypius monachus)雖然傳統上被歸類為鷲鷹科,但在某些分類系統中被納入隼形目。這種大型猛禽主要以腐肉為食,在生態系統中扮演重要角色。
這些隼形目鳥類雖然在外形和生活習性上各有特點,但都展現出一些共同的特徵。它們普遍具有鉤狀的喙,用於撕裂獵物;強壯的爪,用於抓握獵物;銳利的視力,有助於從高空發現獵物。隼形目鳥類的翅膀通常尖銳而有力,適合高速飛行和靈活轉向。許多種類都表現出明顯的性二型現象,雄性通常比雌性體型小,但羽色更鮮艷。
隼形目鳥類在生態系統中扮演著重要的角色,作為頂級掠食者,它們調節獵物種群的數量,維持生態平衡。同時,由於它們對環境變化敏感,常被用作生態系統健康狀況的指示物種。然而,由於棲息地喪失、環境污染和非法捕獵等因素,許多隼形目鳥類的種群正面臨威脅。因此,加強對這些鳥類及其棲息地的保護,不僅對維護生物多樣性至關重要,也關係到整個生態系統的穩定。
隼形目鳥類與人類的關係由來已久。在古代,一些隼類被馴化用於狩獵,形成了獵鷹運動的傳統。現代,這些鳥類在生態旅遊和環境教育中發揮著重要作用,幫助人們瞭解自然界的奧妙和生態保護的重要性。隼形目鳥類的研究不僅涉及生物學和生態學,還為航空技術的發展提供了靈感,體現了自然界與人類科技的緊密聯繫。
佛法僧目
佛法僧目(Coraciiformes)是鳥類分類中一個獨特而多樣的群體,其命名源自希臘文"korax",意為"烏鴉"。這個名稱可能是基於某些佛法僧目鳥類的外形或叫聲與烏鴉相似而得來。然而,這個目中的鳥類實際上與烏鴉並無直接關係,而是包括了一系列色彩豐富、形態各異的鳥類。
佛法僧目鳥類分佈廣泛,遍及世界各大洲,尤其在熱帶和亞熱帶地區種類豐富。這個目包括了許多引人注目的鳥類,如翠鳥、佛法僧、蜂虎等。它們通常具有鮮艷的羽毛、粗壯的喙和相對短小的腿,適應了各種生態環境,從森林到開闊地區,甚至包括水邊棲息地。
普通翠鳥(Alcedo atthis)是一種小型而色彩鮮艷的鳥類,上體呈亮麗的藍色,下體為橙紅色。它們常見於溪流和湖泊邊,以小魚和水生昆蟲為食。翠鳥有著獨特的捕魚方式,常常在水面上方停留,然後迅速俯衝入水捕捉獵物。
三趾翠鳥(Ceyx erithaca)是一種體型更小的翠鳥,以其鮮艷的橙紅色和藍色羽毛而聞名。如其名所示,這種鳥只有三個腳趾,這是它們的一個獨特特徵。
冠翠鳥(Megaceryle lugubris)是一種大型翠鳥,頭上有明顯的冠羽。它們常見於山區溪流,以魚類為主要食物,同時也捕食小型兩棲動物。
歐洲佛法僧(Coracias garrulus)是一種中型鳥類,以其亮麗的藍綠色羽毛而著稱。它們常見於開闊的農田和草地,以大型昆蟲和小型脊椎動物為食。
寬嘴鳥(Eurystomus orientalis)屬於佛法僧科,但與典型的佛法僧有所不同。它們有著寬大的喙和較長的翅膀,常在黃昏時分捕食飛行昆蟲。
綠鬚蜂虎(Merops orientalis)是一種小型而優雅的鳥類,以其綠色羽毛和延長的中央尾羽而聞名。如其名所示,蜂虎主要以蜂類和其他飛行昆蟲為食。
藍喉蜂虎(Merops viridis)比綠鬚蜂虎稍大,有著鮮艷的藍色喉部和綠色體羽。它們常成群在空中捕捉昆蟲,飛行技巧相當精湛。
棕胸佛法僧(Coracias caudatus)是非洲常見的佛法僧物種(種),以其淡藍色的羽毛和棕色的胸部為特徵。它們常棲息在樹枝上,等待地面上的獵物出現。
白胸翡翠(Halcyon smyrnensis)是一種大型翠鳥,但並不局限於水邊棲息。它們有著鮮艷的藍色羽毛和明顯的白色胸部,常見於各種開闊環境。
犀鳥(Buceros bicornis)雖然在某些分類系統中被單獨列為犀鳥目,但傳統上也被歸類為佛法僧目的一部分。這種大型鳥類以其巨大的喙和頭上的角狀突起而聞名。
戴勝(Upupa epops)是一種獨特的鳥類,有著醒目的羽冠和黑白相間的翅膀。它們常在地面上覓食,主要捕食昆蟲和小型無脊椎動物。
綠姬鳥(Todus todus)是加勒比地區的特有物種,體型很小,羽毛鮮綠,喙部扁平而長。它們主要以昆蟲為食,常在低矮植被中活動。
巨翠鳥(Megaceryle maxima)是翠鳥家族中體型最大的成員之一,主要分佈在非洲。它們不僅捕食魚類,還會獵捕小型哺乳動物和爬行動物。
紅腹犀鳥(Aceros nipalensis)是一種中型犀鳥,主要棲息在東南亞的熱帶森林中。它們以其鮮豔的紅色腹部和獨特的叫聲而聞名。
藍胸佛法僧(Coracias cyanogaster)是非洲西部的特有種,有著鮮艷的藍色胸部和腹部,背部則呈現褐色。它們常在開闊的草原和疏林地帶活動。
這些佛法僧目鳥類雖然在外形和生活習性上各有特點,但都展現出一些共同的特徵。它們大多具有鮮艷的羽毛,這不僅有助於種內識別,也在求偶過程中起重要作用。許多佛法僧目鳥類有著強壯的喙,適應了它們的食性,無論是捕魚、捉蟲還是取食果實。此外,這些鳥類普遍具有出色的視力,有助於它們精確地捕捉獵物。
佛法僧目鳥類在生態系統中扮演著重要角色。作為昆蟲和小型脊椎動物的捕食者,它們協助控制這些生物的數量,維持生態平衡。一些種類,如犀鳥,還在種子傳播中發揮重要作用,有助於森林的更新和擴展。然而,由於棲息地喪失、環境污染等因素,許多佛法僧目鳥類正面臨生存威脅。保護這些鳥類及其棲息地,不僅對維護生物多樣性至關重要,也有利於整個生態系統的健康。
佛法僧目鳥類因其獨特的外形和行為,在人類文化中常佔有重要地位。例如,翠鳥在許多文化中被視為美麗和敏捷的象徵,而犀鳥在一些東南亞文化中被視為神聖的鳥類。這些鳥類的研究不僅豐富了我們對生物多樣性的認識,也為生態保護和環境教育提供了重要素材。
鴕鳥目
鴕鳥目(Struthioniformes)的命名源自古希臘語"strouthos",意為"麻雀"或"鳥"。這個命名可能源於早期歐洲人對這些大型非飛行鳥類的誤解或比喻。鴕鳥目包括了世界上最大的現存鳥類,它們都是不能飛行的陸生鳥類,主要分佈在非洲、澳大利亞和南美洲。
然而,需要注意的是,鴕鳥目的分類一直是鳥類學界討論的熱點。
非洲鴕鳥(Struthio camelus)是世界上最大的現存鳥類,也是鴕鳥目的代表性物種。成年雄性可達2.75米高,重達150公斤。它們有長而強壯的腿,能夠高速奔跑,速度可達每小時70公里。非洲鴕鳥的羽毛黑白分明,雄鳥以黑色為主,雌鳥則為灰褐色。
南美洲鴕(Rhea americana)是南美洲的特有種,體型比非洲鴕鳥小,但仍是該地區最大的鳥類。它們的羽毛呈灰褐色,頸部較長,腿部強壯,善於奔跑。美洲鴕的翅膀雖然不能飛行,但在奔跑時可以幫助保持平衡。
澳洲鴯鶓(Dromaius novaehollandiae)是澳大利亞的特有種,也是該大陸上最大的鳥類。它們的羽毛呈暗褐色,頭部和頸部幾乎無羽,露出藍色的皮膚。鴯鶓的腿部肌肉發達,能夠快速奔跑和跳躍。
食火雞(Casuarius casuarius)是分佈在新幾內亞和澳大利亞北部的大型鳥類。它們以頭頂的角質冠和鮮艷的藍色和紅色頭部而聞名。食火雞被認為是世界上最危險的鳥類之一,因為它們的腿部有鋒利的爪子,能夠造成嚴重傷害。
suarius dekkeri)是一種已滅絕的食火雞,化石發現於新幾內亞。它們的體型比現存的食火雞種類要小得多。
恐鳥(Dinornis robustus)是新西蘭特有的已滅絕鳥類,是已知最高的鳥類,可能達到3.6米高。它們可能在人類到達新西蘭後不久就滅絕了。
象鳥(Aepyornis maximus)是馬達加斯加島上的已滅絕物種,被認為是有記錄以來最重的鳥類之一,體重可能超過400公斤。它們產下的蛋是已知最大的鳥蛋。
這些鴕鳥目鳥類雖然在外形和生活習性上各有特點,但都展現出一些共同的特徵。它們都是不能飛行的大型地棲鳥類,翅膀退化,但腿部肌肉發達,適合奔跑。多數種類都有相對細長的頸部和小而扁平的頭部。它們的羽毛通常鬆軟,不像飛行鳥類那樣緊密,這可能與它們不需要飛行有關。
鴕鳥目鳥類在生態系統中扮演著獨特的角色。作為大型草食性或雜食性動物,它們對植被結構和種子傳播有重要影響。同時,它們也是許多捕食者的獵物,在食物鏈中佔據重要地位。
雀形目
雀形目(Passeriformes)的命名源自拉丁文"passer",意為"麻雀"。這個名稱反映了麻雀這種常見鳥類在該目中的代表性地位。雀形目是鳥類中最大的目,包含了超過半數的鳥類種類,約有6,000多種。這些鳥類分佈廣泛,適應了各種生態環境,從熱帶雨林到極地苔原都能找到它們的身影。
雀形目鳥類的共同特徵包括有三個前趾和一個後趾,這種排列使它們能夠牢固地抓住樹枝。多數雀形目鳥類能夠發出複雜的鳴叫,這與它們發達的鳴管有關。此外,許多雀形目鳥類具有性二型現象,即雌雄外形存在明顯差異,通常雄鳥的羽色更為鮮艷。
以下介紹15種常見的雀形目鳥類及其主要特徵:
家麻雀(Passer domesticus)是一種小型雀形目鳥類,全球分佈廣泛。雄性有黑色喉斑和灰色頭頂,雌性則整體呈褐色。它們常見於人類聚居地,適應性極強。
歐洲知更鳥(Erithacus rubecula)以其鮮艷的橙紅色胸部而聞名。這種小型鳥類在歐洲廣泛分佈,常被視為春天的使者。
大山雀(Parus major)是一種體型稍大的山雀,頭部黑色,臉頰白色,胸部有明顯的黑色縱紋。它們常見於woodland(林地)和花園中,以昆蟲和種子為食。
黃鶯(Oriolus chinensis)是一種美麗的中型鳥類,雄性全身金黃色,翅膀和尾部有黑色斑紋。它們的叫聲悅耳,常被稱為"森林笛手"。
夜鶯(Luscinia megarhynchos)以其美妙的歌聲而聞名,尤其在夜間更為動聽。它們的外形較為樸素,全身褐色,但其歌喉使它們成為歐洲最受歡迎的鳥類之一。
燕子(Hirundo rustica)以其長而分叉的尾巴和優雅的飛行姿態而聞名。它們的上體呈藍黑色,具有金屬光澤,下體為白色,有時帶有淡褐色。燕子是候鳥,每年往返於繁殖地和越冬地之間。
烏鴉(Corvus corone)是一種大型的全黑色鳥類,以其智慧和適應能力而聞名。它們的叫聲低沉嘶啞,在許多文化中都有重要的象徵意義。
畫眉(Garrulax canorus)是一種中型鳥類,羽毛以褐色為主,頭部和胸部有明顯的斑紋。它們以優美的歌聲而聞名,在中國文化中佔有重要地位。
藍鵲(Urocissa erythrorhyncha)是一種大型的美麗鳥類,全身藍色,頭部黑色,尾巴長而分層。它們常見於東亞的森林地區,具有高度的智力和社交能力。
八哥(Acridotheres cristatellus)是一種中型鳥類,全身黑色,翅膀上有白色斑塊。它們常見於城市和農村地區,有模仿各種聲音的能力。
百靈鳥(Alauda arvensis)以其高空盤旋時發出的悅耳歌聲而聞名。它們的羽毛呈褐色帶有斑點,能夠很好地融入草地環境。
翠鳥(Alcedo atthis)雖然不屬於雀形目,但因其普遍性常被一起討論。它們以鮮艷的藍色上體和橙色下體而聞名,常見於水邊,以小魚為食。
雲雀(Alauda gulgula)是一種小型地棲鳥類,羽毛呈褐色帶有斑點。它們以高空飛行時發出的連續鳴叫而聞名,常見於開闊的草原和農田。
這些雀形目鳥類展現了該目的多樣性和適應性。它們在生態系統中扮演著重要角色,不僅是許多食物鏈的重要一環,也是生態平衡的指標。例如,許多雀形目鳥類以昆蟲為食,幫助控制害蟲數量;而一些種子食性的鳥類則在植物種子的傳播中發揮重要作用。
雀形目鳥類與人類的關係密切。許多種類因其悅耳的歌聲或美麗的羽毛而受到人們的喜愛,成為觀鳥愛好者和自然愛好者關注的焦點。在文學、藝術和民間傳說中,雀形目鳥類常常扮演重要角色,反映了人類對自然界的認知和情感。
雀形目
雀形目(Passeriformes)鳥類中最大的目,包含了眾多的科。以下介紹雀形目中最常見的科,以及每個科的代表性鳥類和特徵:
雀科(Passeridae)的代表性鳥類是家麻雀。雀科鳥類通常體型小,喙短而粗壯,適合取食種子。家麻雀(Passer domesticus)適應性極強,全球分佈廣泛。山麻雀稀有保護種。
椋鳥科(Sturnidae),羽毛常有金屬光澤。多群居,鳴聲多樣。適應性強,常見於開闊地區和城市。部分種類具模仿他鳥叫聲的能力。全球約110種,分布廣泛。冠八哥(全身黑、有黑剛毛如冠)、白尾八哥、家八哥(褐羽、黃眼眶)、黑領椋鳥、歐洲椋鳥。
鴉科(Corvidae)是一個廣泛分佈的科,其代表性鳥類包括烏鴉、藍鵲、樹鵲、喜鵲和松鴉。鴉科鳥類通常體型較大,智力高,社交能力強。它們的喙粗壯,適合多樣化的飲食。烏鴉(Corvus corax)全身黑色,有光澤,以其聰明和適應性強而聞名。
燕科(Hirundinidae)的代表性鳥類是家燕。燕科鳥類以其流線型的身體、分叉的尾巴和優雅的飛行而著稱。它們主要以飛蟲為食,常在空中捕食。常見的有家燕、赤腰燕、洋燕、毛腳燕、棕沙燕、灰沙燕。
鶯科(Sylviidae)包括了許多小型鳴禽,如柳鶯和蘆鶯。鶯科鳥類通常體型小巧,羽毛顏色不鮮艷,但歌聲優美。台灣常見夜鶯、。
山雀科(Paridae)的代表性鳥類包括大山雀和藍山雀。山雀科鳥類通常體型小,活潑好動,善於攀爬。它們的喙短而有力,適合取食種子和昆蟲。台灣常見黃山雀、青背山雀、媒山雀。
鶇科(Turdidae)包括知更鳥和烏鶇等。鶇科鳥類通常體型中等,腿長,適合在地面覓食。許多種類都有優美的歌聲。台灣常見藍磯鶇、虎鶇、白尾鴝、
鵯科(Pycnonotidae) 台灣常見紅嘴黑鵯、白頭翁、烏頭翁、白環鸚嘴鵯
文鳥科(Estrildidae)包括了許多小型的雀形目鳥類,如珍珠鳥和斑胸鳩。文鳥科鳥類通常體型小巧,喙短而粗,主要以種子為食。常見的白腰文鳥、斑文鳥、黑頭文鳥。
鶺鴒科(Motacillidae),台灣常見白鶺鴒、灰鶺鴒、山鶺鴒
鶲科(Muscicapidae)包括許多以捕食飛蟲為生的小型鳥類,如鶲和鴝。鶲科鳥類通常有扁平的喙和發達的飛行肌肉,善於在空中捕捉昆蟲。台灣常見小剪尾、鉛色水鶇、紫嘯鶇、藍磯鶇、白腰鵲鴝、黃尾鴝、藍歌鴝、鵲鴝、白眉地鶇、黃腹琉璃、虎斑地鶇。
百靈科(Alaudidae)的代表性鳥類是雲雀。百靈科鳥類通常棲息在開闊地帶,有發達的鳴唱能力。雲雀(Alauda arvensis)以其高空盤旋時發出的悅耳歌聲而聞名,常見於草原和農田。
畫眉科 Timaliidae,台灣常見山紅頭、大彎嘴、小彎嘴
鸚嘴科(Psittaculidae)雖然大多數鸚鵡不屬於雀形目,但鸚嘴科是雀形目中的例外。代表性鳥類包括粉紅鸚嘴。鸚嘴科鳥類通常有彎曲的喙和鮮艷的羽毛。。
這些科的鳥類展現了雀形目的多樣性,從體型、羽色到生活習性都有很大差異。它們適應了各種生態環境,在生態系統中扮演著重要角色。有些是重要的授粉者和種子傳播者,有些則通過捕食昆蟲來控制害蟲數量。同時,許多雀形目鳥類因其優美的歌聲或鮮艷的羽毛而受到人類的喜愛,在文化和藝術中佔有重要地位。
鸊鷉目
鸊鷉目(Podicipediformes)的命名源自拉丁文"podex"(臀部)和"pes"(腳),意指這些鳥類的腿部位於身體後端靠近臀部的位置。這個特徵反映了鸊鷉目鳥類對水生環境的適應。鸊鷉目是一個相對較小的目,包含了一些高度適應水生生活的鳥類。這些鳥類以其出色的潛水能力和獨特的外形而聞名。
鸊鷉目鳥類的共同特徵包括流線型的身體,有利於水中游泳;腿部位於身體後端,使它們能夠有力地推動身體在水中前進;腳趾有蹼相連,但不同於其他水鳥,鸊鷉的腳趾是分瓣的,每個趾都有獨立的蹼。它們的喙通常細長而尖銳,適合捕捉魚類和水生昆蟲。鸊鷉目鳥類在陸地上行動笨拙,但在水中卻非常靈活,能夠快速潛水和游泳。
鸌形目
鸌形目(Procellariiformes)的命名源自拉丁文"procella",意為"暴風雨"。這個名稱反映了鸌形目鳥類常常在海洋上空飛翔,即使在惡劣的天氣條件下也能夠生存。鸌形目是一個適應海洋生活的鳥類目,包括了信天翁、海燕、潛水鸌等海鳥。這些鳥類以其出色的飛行能力和對海洋環境的適應而聞名。
鸌形目鳥類的共同特徵包括管狀的鼻孔,這種結構有助於排出體內多餘的鹽分;強壯的鉤狀喙,適合捕捉魚類和魷魚;腳蹼發達,有利於在水面上活動。大多數鸌形目鳥類具有長而窄的翅膀,能夠在海面上方長時間滑翔。它們通常具有防水的羽毛,能夠適應長期在海上生活的需求。
鴿形目
鴿形目(Columbiformes)的命名源自拉丁文"columba",意為"鴿子"。這個名稱反映了鴿子在該目中的代表性地位。鴿形目是一個中等規模的鳥類目,包括了鴿子和鳩等常見鳥類。這些鳥類以其獨特的外形、溫和的性格和廣泛的分佈而聞名。
鴿形目鳥類的共同特徵包括圓潤的身體,相對較小的頭部,短而纖細的喙,喙基部常有蠟膜。它們的腳短而有力,適合在地面行走和棲息在樹枝上。鴿形目鳥類通常有發達的嗉囊,用於儲存和軟化食物。它們的羽毛柔軟,常具有金屬光澤。鴿形目鳥類以其獨特的"咕咕"叫聲而聞名,這種聲音在求偶和領域宣示中起重要作用。以下介紹常見的鴿形目鳥類及其主要特徵:
1. 岩鴿(Columba livia)是最廣為人知的鴿子種類,也是家鴿的祖先。野生岩鴿通常呈灰色,翅膀上有兩道黑色橫帶。它們原產於歐亞大陸和北非,但現在已經在世界各地廣泛分佈。岩鴿適應性極強,能夠在城市環境中生存繁衍。
2. 綠翅鳩(Chalcophaps indica)是一種美麗的小型鴿子,主要分佈在東南亞和大洋洲。它們的背部呈現鮮艷的綠色金屬光澤,翅膀上有白色斑塊。綠翅鳩常在森林地面覓食,飛行時發出響亮的拍翅聲。
3. 斑鳩(Streptopelia chinensis)廣泛分佈於亞洲。它們的羽毛呈淡褐色,頸部有特徵性的黑白相間的斑紋。斑鳩常見於城市和鄉村地區,以其溫和的性格和優雅的外形而受歡迎。
4. 珠頸斑鳩 (Spilopelia chinensis): 中小型鳩鴿,頸部有黑白相間的斑紋。常見於城市和鄉村地區,以種子和穀物為食。叫聲低沉而有節奏。
5. 金背鳩 (Streptopelia orientalis): 體型較大的鳩鴿,背部呈金褐色。棲息於森林和開闊地帶,食物包括種子、漿果和小昆蟲。叫聲悅耳動聽。
6. 灰林鳩 (Columba pulchricollis): 中型林鳩,全身灰色,頸部有白色半環。主要棲息於山區森林,喜歡在樹上築巢。以果實和種子為主食。
7. 紅頭綠鳩 (Treron formosae): 體型適中,頭部紅色,身體大部分為綠色。主要棲息於低海拔森林,常成群活動。以各種果實為食。銀鳩(Zenaida macroura)廣泛分佈於北美洲。它們的羽毛呈淡褐色,尾部有特徵性的白邊。銀鳩以其悅耳的鳴叫聲而聞名。
8. 綠鳩(Treron)是一類廣泛分佈於亞洲和非洲的鴿子。它們的羽毛主要呈現綠色,能夠很好地隱藏在樹冠中。
雁形目
雁形目(Anseriformes)的命名源自拉丁文"anser",意為"雁"。這個名稱反映了雁在該目中的代表性地位。雁形目是一個包含了鴨、鵝、天鵝等水禽的鳥類目。這些鳥類以其水生適應性和獨特的外形特徵而聞名。
雁形目鳥類的共同特徵包括扁平的喙,喙端有特殊的構造稱為"篩板",用於過濾水中的食物;腳蹼發達,適合在水中游泳;身體通常呈流線型,有利於水中活動;羽毛緻密且有油脂,具有良好的防水性。雁形目鳥類通常具有sexual dimorphism(性二型現象),雄性往往比雌性更為艷麗。常見的雁形目鳥類及其主要特徵如下:
1. 綠頭鴨(Anas platyrhynchos)是最廣為人知的野鴨種類,也是家鴨的祖先。雄性頭部呈現鮮艷的綠色金屬光澤,頸部有白色環,身體灰褐色,尾部上翹。雌性全身褐色帶有斑點,適合偽裝。綠頭鴨廣泛分佈於北半球,適應性極強。
2. 大白鵝(Anser cygnoides)是亞洲原產的大型鵝類,也是家鵝的主要祖先之一。它們體型碩大,頸部修長,喙部粗壯,羽毛以灰褐色為主。大白鵝在野外主要分佈於東亞地區,但馴化品種已遍及全球。
3. 疣鼻天鵝(Cygnus olor)以其優雅的姿態和純白的羽毛而聞名。它們的特徵是喙基部有一個黑色的疣狀突起,頸部修長呈S形彎曲。疣鼻天鵝原產於歐亞大陸,現已被引入北美洲等地。
4. 琵嘴鴨(Spatula cpeata)以其特大而扁平的喙而得名。雄性繁殖羽頭部呈綠色,胸部白色,腹部栗色。這種鴨子的喙特別適合過濾水中的小型生物。
5. 鴛鴦(Aix galericulata)是東亞特有的一種小型鴨類,以其艷麗的羽毛而聞名。雄性有鮮豔的橙色羽扇和紫紅色的胸部,在中國文化中象徵愛情和忠誠。
6. 黑天鵝(Cygnus atratus)是澳大利亞特有的天鵝種類。它們全身羽毛呈黑色,與白色飛羽形成鮮明對比,喙呈鮮紅色。黑天鵝優雅的姿態使其成為澳大利亞的標誌性鳥類之一。
7. 鸊鵜(Mergus merganser)是一種大型潛水鴨,有著鋸齒狀的喙,特別適合捕捉魚類。雄性頭部呈深綠色,身體主要為白色,背部黑色。
8. 灰雁(Anser anser)是歐亞大陸常見的大型雁類,也是家鵝的另一個主要祖先。它們全身灰色,體型碩大,是重要的狩獵對象和觀鳥目標。
這些雁形目鳥類展現了該目的多樣性和適應性。在生態系統中,雁形目鳥類扮演著重要角色,它們不僅是許多捕食者的獵物,也通過採食和排泄影響水生植物的分佈和水體的營養循環。許多雁形目鳥類是長距離遷徙者,其遷徙路線的研究為我們瞭解全球氣候變化和生態系統健康提供了重要線索。
鵜 形目
鵜形目(Pelecaniformes)的命名源自古希臘語「(pelekán),意為「鵜鶘」。這個名稱反映了鵜鶘作為該目中最具代表性的鳥類之一的地位。鵜形目鳥類通常與水域環境密切相關,包括海洋、湖泊、河流和沼澤地等。主要的科有鷺科 (Ardeidae)、鵜鶘科 (Pelecanidae)、鸕鶿科 (Phalacrocoracidae)、鰹鳥科 (Sulidae)、軍艦鳥科 (Fregatidae)、鹮科 (Threskiornithidae)。
這些鳥類在形態和行為上都有一些共同特徵,如腳蹼、長喙,以及擅長游泳和捕魚等。以下是常見的鵜形目鳥類及其特徵描述:
1. 朱鷺(Crested Ibis)是一種瀕危的鵜形目鳥類。它們的羽毛呈粉紅色,頭部有一撮羽冠。朱鷺有著長而彎曲的紅色喙,這是它們的標誌性特徵。由於棲息地破壞和環境污染,朱鷺的數量曾經急劇減少,現在正在進行保護和繁殖計劃。
2. 琵鷺(Spoonbill)是鵜形目中的另一種獨特鳥類。它們最顯著的特徵是扁平而呈匙狀的喙端,這種特殊的喙形使它們能夠在淺水中左右擺動頭部來過濾食物。琵鷺的羽毛通常呈白色或粉紅色。
3. 黃頭鷺(Bubulcus coromandus)屬於「夏候鳥」的一種,是全世界上分佈最廣的鷺科鳥類之一,牠們在台灣雖然終年可見,卻有著在島內遷移的奇特現象,每年秋冬季,北部的黃頭鷺會移居到較溫暖的南部渡冬,來年春天才又回到北台灣繁殖,而目前所見的黃頭鷺,可說是兼具著遷徙與留鳥雙重特性;常見於農村田間、草原、沼澤等地,因常跟隨牛隻或耕耘機附近吃蟲,或棲息於牛背上,故又俗稱為「牛背鷺」。在繁殖季時常與小白鷺、夜鷺等其他鷺科鳥類,共同築巢於竹林、木麻黃林之中 ,成群行動,集體繁殖。其主要特徵為嘴黃腳黑,頸粗短;繁殖期,頭、頸部會出現橘黃色飾羽。
4. 蒼鷺(Grey Heron)是一種大型的鵜形目鳥類。它們的羽毛主要呈灰色,頭部和頸部有黑白相間的條紋。蒼鷺有著長而有力的喙,能夠捕捉魚類和其他小型動物。
5. 大白鷺(Great Egret)是一種優雅的大型鷺鳥。它們全身羽毛潔白,有著修長的脖子和細長的黑色腿。大白鷺的喙呈黃色,在繁殖季節會變成黑色。
6. 小白鷺(Little Egret)與大白鷺相似,但體型較小。它們同樣全身潔白,但腿部為黑色,腳趾呈黃色。小白鷺的喙細長且呈黑色。
7. 夜鷺(Black-crowned Night Heron)是一種中型鷺鳥,活動時間主要在黃昏和夜間。成鳥的頭頂和背部呈黑色,翅膀和尾部為灰色,腹部為白色。夜鷺有著粗壯的喙和相對較短的腿。
8. 黑面麻鷺(Gorsachius melanolophus) 是一種中型鷺鳥,成鳥頭頂和後頸為黑色,有長而蓬鬆的冠羽,面部和喉部為黑色,活動時間主要在黃昏。主要棲息在亞熱帶和熱帶地區的樹林間,獨行地棲性覓食,在樹上築巢,通常選擇離水較近的樹木。
9. 鵜鶘(Pelican)是鵜形目中最為人熟知的成員之一。它們體型龐大,最顯著的特徵是巨大的喙和喉囊。喉囊用於捕魚和儲水,能夠容納大量的魚類。鵜鶘有多個品種,如白鵜鶘和褐鵜鶘,它們的羽毛顏色各不相同,但都具有相似的體型特徵。
10. 鸕鶿(Cormorant)是另一種常見的鵜形目鳥類。它們體型中等,羽毛通常呈黑色或深褐色,具有長而彎曲的頸部和鉤狀的喙。鸕鶿是優秀的潛水者,能夠在水下追捕魚類。一些常見的鸕鶿種類包括大鸕鶿和小鸕鶿。
11. 軍艦鳥(Frigatebird)是鵜形目中的空中霸主。它們有著修長的身體、長而尖的翅膀,以及分叉的尾巴。雄性軍艦鳥在求偶季節會膨脹其鮮紅色的喉囊,這是它們最顯著的特徵之一。軍艦鳥以其出色的飛行能力聞名,能夠長時間在空中滑翔。
12. 鰹鳥(Booby)是一種中型海鳥,屬於鵜形目。它們有著流線型的身體,長而尖的喙,以及楔形的尾巴。鰹鳥以其俯衝捕魚的技能而聞名,能夠從高空直接俯衝入水捕獵。藍面鰹鳥和紅腳鰹鳥是較為常見的品種。
13. 蛇鵜(Anhinga),也被稱為蛇鳥或水火雞,是一種獨特的鵜形目鳥類。它們有著細長的脖子和尖銳的喙,羽毛通常呈黑色或深褐色。蛇鵜善於游泳和潛水,經常可以看到它們只露出脖子和頭部在水面上游動,這種姿態讓它們看起來像一條遊動的蛇,因此得名。
這些鵜形目鳥類雖然在外形和行為上各有特點,但它們都與水域環境有著密切的聯繫。大多數鵜形目鳥類都是出色的捕魚者,擁有適應水生環境的特殊結構,如腳蹼、長喙等。許多種類都能夠進行長距離遷徙,在生態系統中扮演著重要角色。
鶴形目
鶴形目(Gruiformes)的命名源自拉丁文「grus」,意為「鶴」。這個名稱反映了鶴類在該目中的重要地位和代表性。鶴形目包括了一些體型較大的涉禽,如鶴、秧雞和鷓鴣等。這些鳥類通常具有長腿、長頸和長喙的特徵,適應了在濕地、草原和森林等多樣化環境中生活。以下是常見的鶴形目鳥類及其特徵描述:
1. 丹頂鶴是鶴形目中最為人熟知的成員之一。它們體型高大優雅,羽毛以白色為主,頭頂有鮮明的紅色冠羽,這是它們最顯著的特徵。丹頂鶴的翼尖為黑色,形成與白色身體的鮮明對比。它們有著細長的黑色腿和長而尖的喙,非常適合在沼澤地和淺水區覓食。
2. 白鶴是另一種優雅的大型鶴類。如其名所示,它們全身羽毛潔白,只有臉部周圍有一小片裸露的紅色皮膚。白鶴的體型略小於丹頂鶴,但同樣具有修長的頸部和腿。它們主要棲息在凍原和濕地環境中,是極其珍稀的鳥類。
3. 秧雞是鶴形目中體型較小的成員。它們的身體扁平,適合在密集的植被中穿行。秧雞的羽毛通常呈褐色或灰色,有助於在沼澤和蘆葦叢中隱蔽。它們有著較短的喙和強壯的腿,能夠靈活地在水陸之間活動。
4. 水雉是一種獨特的鶴形目鳥類。它們最顯著的特徵是極長的腳趾,這使得它們能夠在水生植物的葉子上行走。水雉的羽毛在繁殖期和非繁殖期有明顯的變化,繁殖期的雄鳥有著黑色的頭部和金黃色的背部,非常引人注目。
5. 董雞是一種小型的鶴形目鳥類。它們的羽毛呈褐色,有著黑白相間的條紋,非常適合在蘆葦叢中隱蔽。董雞的喙較短,但很堅固,適合啄食植物和小型動物。
6. 鷓鴣是鶴形目中的一個獨特群體。它們的體型較小,羽毛通常呈褐色或灰色,有複雜的斑點和條紋圖案。鷓鴣的腿相對較短,但非常有力,適合在草地和灌木叢中奔跑。
7. 白腹秧雞是一種中型秧雞。如其名所示,它們的腹部呈白色,背部和翅膀呈棕色或灰褐色。白腹秧雞有著紅色的喙和腿,這與它們的羽毛形成鮮明對比。它們主要棲息在濕地和沼澤地區。
8. 鴇是鶴形目中的另一個獨特群體,有時被單獨分類為鴇形目。它們是大型的地棲鳥類,主要棲息在開闊的草原和荒漠環境中。鴇的羽毛通常呈褐色或灰色,有複雜的斑點和條紋圖案,有助於偽裝。雄性鴇在求偶時會進行誇張的展示行為,膨脹喉囊並豎起羽毛。
這些鶴形目鳥類雖然在形態和行為上各有特點,但它們都與特定的生態環境有著密切的聯繫。許多鶴形目鳥類都適應了濕地或草原環境,擁有長腿和長頸等特徵,有利於在這些環境中覓食和生存。一些種類,如鶴,以其優雅的外形和複雜的求偶舞蹈而聞名。
3.7a 台灣鳥類的棲息環境
台灣擁有豐富多樣的地理環境和生態系統,為眾多鳥類提供了理想的棲息地。從海岸線到高山,從平原到森林,每一種環境都孕育著獨特的鳥類群落。
首先,讓我們從海岸線開始談起。台灣四面環海,擁有長達1,566公里的海岸線,包括沙灘、礁岩、紅樹林和河口濕地等多樣化的棲地。這些地方是許多水鳥和候鳥的重要棲息地。在沙灘和潮間帶,我們常能看到鷸科(Scolopacidae)和鴴科(Charadriidae)的鳥類,如東方環頸鴴和黑腹濱鷸。這些鳥類以沙灘上的小型無脊椎動物為食,長而纖細的喙正是為了在沙中捕食而進化的。在紅樹林和河口濕地,則能發現如鷺科(Ardeidae)的鳥類,包括大白鷺和蒼鷺。這些地方不僅為鳥類提供食物,也是許多候鳥遷徙途中的重要中繼站。
向內陸移動,我們來到了平原和低海拔丘陵地帶。這裡的農田、草地和灌叢是許多陸鳥的家園。在稻田和農地周圍,常見的鳥類包括麻雀(Passer montanus)和白頭翁(Pycnonotus sinensis)。這些鳥類已經很好地適應了人類活動,能在農業環境中覓食和繁殖。在較為開闊的草地,我們可能會遇到百靈科(Alaudidae)的鳥類,如小雲雀。這些鳥類通常以地面覓食,並在地上築巢。
隨著海拔的升高,我們進入了森林地帶。台灣的森林覆蓋率高達58.5%,為眾多鳥類提供了廣闊的生存空間。在低海拔的闊葉林中,我們可以發現如五色鳥(Megalaima nuchalis)和竹雞(Bambusicola thoracicus)等鳥類。五色鳥以其鮮艷的羽毛和獨特的叫聲聞名,常在樹冠層活動。竹雞則偏好在地面和灌木叢中活動,以各種植物種子和小型無脊椎動物為食。
中海拔的混合林是台灣特有種鳥類的重要棲息地。例如,台灣藍鵲(Urocissa caerulea)就主要分佈在海拔300到1,200米的森林中。這種美麗的鳥類不僅是台灣的特有種,也是台灣的國鳥之一。牠們通常成群活動,以各種果實、昆蟲和小型脊椎動物為食。在這個海拔段,我們還能發現如台灣噪眉(Alcippe morrisonia)和繡眼畫眉(Alcippe brunnea)等小型鳴禽。這些鳥類主要在樹冠下層和灌木叢中活動,以昆蟲為主要食物來源。
再往上,進入高海拔的針葉林和高山草原,我們會遇到一些適應寒冷氣候的鳥類。例如,深山竹雞(Arborophila crudigularis)是台灣特有種,主要生活在海拔1,200到3,000米的森林中。這種鳥類喜歡在陰暗潮濕的林下活動,以各種植物種子、漿果和昆蟲為食。在高山草原,我們可能會看到花嘴鴉(Pyrrhocorax pyrrhocorax)。這種鳥類適應了高海拔的環境,常在開闊的草地上覓食。
台灣的高山地區還有一些特別的微棲地,如高山湖泊和溪流。這些地方為一些特殊的鳥類提供了棲息環境。例如,在高山溪流邊,我們可能會遇見河烏(Cinclus pallasii)。這種鳥類有著獨特的生活方式,能夠在湍急的溪流中潛水捕食水生昆蟲。
值得一提的是,台灣的地理位置使它成為許多候鳥的重要中繼站和越冬地。每年秋冬季節,大量的候鳥會從北方遷徙到台灣。其中,鰲鼓濕地和四草濕地等地方是重要的水鳥棲息地。在這些地方,我們可以看到大批的雁鴨科(Anatidae)鳥類,如小水鴨和綠頭鴨。這些鳥類利用台灣溫暖的冬季氣候和豐富的食物資源來度過寒冷的冬天。
台灣的地形複雜,從海平面到海拔近4,000米的高山,在短距離內就能經歷多種氣候帶,這為鳥類提供了多樣化的棲息環境。然而,隨著人類活動的擴張,許多鳥類的棲息地正面臨威脅。城市化導致的棲地破碎化和污染,以及氣候變化帶來的影響,都對鳥類的生存構成了挑戰。
為了保護這些珍貴的鳥類資源,台灣設立了多個國家公園和保護區。例如,墾丁國家公園不僅保護了珍貴的海岸生態系統,也是許多候鳥的重要棲息地。玉山國家公園則保護了大面積的高山生態系統,為許多高海拔鳥類提供了安全的家園。此外,台灣還設立了許多野鳥保護區,如關渡自然公園,專門為保護鳥類棲息地而設立。
除了設立保護區,台灣還採取了多項措施來保護鳥類棲息地。例如,推廣友善耕作,減少農藥使用,以保護在農田中覓食的鳥類。在城市規劃中,也越來越多地考慮到綠地和水域的保留,為城市鳥類提供棲息空間。此外,針對一些瀕危鳥類,如黑面琵鷺,還制定了專門的保護計劃,包括棲地修復和繁殖地保護等措施。
台灣豐富的鳥類資源不僅具有重要的生態價值,也為觀鳥愛好者提供了絕佳的觀察機會。每年都有大量的觀鳥者來到台灣,尤其是在候鳥遷徙的季節。這不僅促進了生態旅遊的發展,也提高了公眾對鳥類保護的認識。許多學校和社區組織也開展了鳥類教育項目,讓更多人瞭解鳥類及其棲息環境的重要性。
3.8 台灣常見的候鳥
台灣位處東亞-澳大利西亞遷徙路線上的重要節點,每年都有大量候鳥途經或在此停留。這些候鳥的到來為台灣的生態系統帶來了季節性的變化和活力。讓我們深入探討台灣常見的候鳥,包括它們的季節性出現、種類、特徵以及珍稀程度。
秋季是候鳥遷徙的開始。通常從九月下旬開始,越來越多的候鳥會陸續抵達台灣。其中,水鳥是最引人注目的群體之一。雁鴨科(Anatidae)的鳥類如小水鴨(Anas crecca)和綠頭鴨(Anas platyrhynchos)是常見的冬候鳥。小水鴨體型嬌小,雄鳥頭部有著明顯的綠色羽毛,而綠頭鴨則以雄鳥亮麗的綠色頭部著稱。這些鴨子通常會在台灣的濕地和水域度過整個冬天,直到次年春天北返繁殖地。雖然這些鴨子在全球範圍內並不稀有,但它們大量聚集的景象仍然十分壯觀。
除了雁鴨科,鷸鴴科(Charadriidae)的鳥類也是重要的候鳥群體。東方環頸鴴(Charadrius alexandrinus)和黑腹濱鷸(Calidris alpina)等都是常見的過境鳥或冬候鳥。這些鳥類通常在海岸線、濕地或河口地區覓食,以各種無脊椎動物為食。牠們纖細的身形和長喙是為了在泥灘和沙灘上覓食而演化的。雖然這些鳥類個體較小,但它們往往成群出現,形成令人印象深刻的景象。
在鷺科(Ardeidae)鳥類中,大白鷺(Ardea alba)和中白鷺(Egretta intermedia)是常見的冬候鳥。這些優雅的大型水鳥以其純白的羽毛和修長的頸部聞名。牠們常在淺水區域覓食,捕捉魚類和其他水生生物。雖然這些鷺鳥在全球範圍內分佈廣泛,但它們在台灣的大量聚集仍然是冬季濕地的一大特色。
其中最為珍稀的水鳥當屬黑面琵鷺(Platalea minor)。這種全球瀕危物種主要在東亞繁殖,冬季會南下到包括台灣在內的東南亞地區越冬。黑面琵鷺以其獨特的勺狀喙和黑色臉部著稱,是許多觀鳥者和保育人士關注的焦點。台灣的四草濕地和七股濕地是黑面琵鷺重要的越冬地,每年到訪的數量約占全球總數的一半左右,因此台灣在這個物種的保育中扮演著關鍵角色。
除了水鳥,許多陸鳥也是台灣重要的候鳥。其中,令人矚目的是猛禽類。每年秋季,大批猛禽會途經台灣,其中最為人知的是灰面鵟鷹(Butastur indicus)。這種中型猛禽以其灰色的面部和褐色的體羽為特徵,每年九月到十月間會大量出現在台灣的天空中,形成壯觀的遷徙景象。墾丁國家公園的龍磐草原是觀察這種鳥類遷徙的最佳地點之一。雖然灰面鵟鷹並不是全球性的瀕危物種,但其大規模遷徙的景象仍然十分珍貴。
另一種引人注目的候鳥是紅尾伯勞(Lanius cristatus)。這種小型猛禽以其褐色的羽毛和黑色眼罩狀斑紋為特徵,通常在秋季抵達台灣,在開闊的草地和農田地區越冬。紅尾伯勞雖然體型不大,但性情兇猛,常常可以看到牠們將獵物插在荊棘上的行為,這也讓牠們贏得了「屠夫鳥」的綽號。
在鳴禽中,燕科(Hirundinidae)的鳥類如家燕(Hirundo rustica)和赤腰燕(Cecropis daurica)都是常見的夏候鳥。這些敏捷的飛行者通常在春季抵達台灣繁殖,秋季再南返越冬。牠們優雅的飛行姿態和叉狀的尾部是其顯著特徵。雖然這些燕子在全球範圍內分佈廣泛,但它們在城市和鄉村地區築巢繁殖的景象仍然是台灣夏季的一大特色。
另一種值得關注的鳴禽是黃眉柳鶯(Phylloscopus inornatus)。這種小型鳥類以其黃綠色的羽毛和明顯的眉斑為特徵,是台灣常見的冬候鳥。牠們通常在秋季抵達,在台灣的森林和公園中度過冬天。雖然個體較小,但其清脆的鳴叫聲常能引起觀鳥者的注意。
在鶇科(Turdidae)鳥類中,白腹鶇(Turdus pallidus)和赤腹鶇(Turdus chrysolaus)都是台灣常見的冬候鳥。這些鳥類通常體型較大,以果實和無脊椎動物為食。白腹鶇以其灰褐色的上體和白色的腹部為特徵,而赤腹鶇則以其橙紅色的腹部引人注目。這些鶇類雖然在原產地並不稀有,但它們的到來為台灣的冬季帶來了豐富的色彩和生機。
有些候鳥雖然不常見,但卻極為珍稀。例如,黑嘴端鳳頭燕鷗(Thalasseus bernsteini)是全球極度瀕危的物種,全球數量可能不足3,000隻。每年都有少量個體在台灣的海岸地區被觀察到,這使得台灣在該物種的保育中扮演著重要角色。
3.9 台灣常見的留鳥
台灣地處亞熱帶,擁有豐富多樣的生態環境,為眾多鳥類提供了理想的棲息地。在台灣的鳥類中,留鳥是一個重要的組成部分。這些鳥類終年生活在台灣,不進行長距離遷徙,因此成為台灣生態系統中穩定且重要的成員。
在城市和鄉村地區常見的留鳥。麻雀(Passer montanus)無疑是最為人熟知的一種。這種小型鳥類已經很好地適應了人類環境,幾乎在台灣的每個角落都能看到它們的身影。麻雀體型小巧,羽毛以褐色為主,雄性和雌性外表相似。它們常常成群出現,在地面覓食或在建築物上築巢。麻雀的適應能力極強,能夠利用各種人造結構築巢,這使得它們在城市環境中也能茁壯成長。
另一種在人類聚居地常見的留鳥是白頭翁(Pycnonotus sinensis)。這種中型鳥類以其特徵性的白色頭部和黑色冠羽而聞名。白頭翁的叫聲悅耳動聽,常在清晨和黃昏時分歌唱。它們主要以果實和昆蟲為食,因此在果園和公園中經常可以看到它們的身影。白頭翁適應性強,在城市和鄉村地區都能生存良好,是台灣最常見的鳥類之一。
在較為開闊的環境中,如農田和草地,我們常能看到喜鵲(Pica pica)的身影。這種黑白相間的大型鳥類以其長尾和金屬光澤的羽毛而引人注目。喜鵲是知名的智慧鳥類,它們能夠使用工具,還具有自我認知的能力。在台灣,喜鵲常在電線桿或大樹上築巢,其巨大的球形巢很容易被人發現。
樹林和公園中,綠繡眼(Zosterops japonicus)是一種常見的小型留鳥。這種鳥類以其鮮艷的黃綠色羽毛和明顯的白色眼圈為特徵。綠繡眼通常成小群活動,它們敏捷靈活,常在樹枝間跳躍覓食。除了昆蟲外,綠繡眼還喜歡吸食花蜜,因此在開花的季節,我們常能看到它們在花叢中忙碌的身影。
在較為茂密的森林中,台灣竹雞(Bambusicola sonorivox)是一種常見的地棲性留鳥。這種鳥類是台灣的特有種,主要分佈在低海拔至中海拔的森林和竹林中。台灣竹雞體型圓潤,羽毛以褐色為主,有著複雜的斑紋。它們通常在地面活動,以各種植物種子和小型無脊椎動物為食。雖然台灣竹雞較為害羞,不易被直接觀察到,但它們響亮的叫聲常能在森林中聽到。
山區常見的留鳥中,台灣藍鵲(Urocissa caerulea)無疑是最引人注目的一種。這種美麗的鳥類不僅是台灣的特有種,也是台灣的國鳥之一。台灣藍鵲體型較大,羽毛以亮麗的藍色為主,長長的尾巴末端有白色斑點。它們通常成群活動,主要分佈在海拔300到1,200米的森林中。台灣藍鵲聰明機智,食性廣泛,從果實到小型脊椎動物都在其食譜之中。
在中低海拔的森林中,五色鳥(Megalaima nuchalis)是一種常見且引人注目的留鳥。這種鳥類以其鮮艷的羽毛而聞名,頭部和頸部有紅、黃、藍、綠、黑五種顏色,故而得名。五色鳥主要以果實為食,特別喜歡無花果。它們通常在樹冠層活動,用強壯的喙在樹幹上鑿洞築巢。五色鳥的叫聲低沉有力,在繁殖季節尤為常見。
另一種在森林中常見的留鳥是大彎嘴(Pomatorhinus erythrocnemis)。這種鳥類是台灣的特有亞種,以其獨特的下彎長喙為特徵。大彎嘴主要生活在低海拔至中海拔的森林中,常在地面或灌木叢中覓食。它們通常成對或小群活動,以各種昆蟲和植物種子為食。大彎嘴的叫聲清亮悅耳,是森林中常聽到的聲音之一。
在較高海拔的森林中,冠羽畫眉(Yuhina brunneiceps)是一種常見的留鳥。這種小型鳥類是台灣的特有種,以其特徵性的直立冠羽而聞名。冠羽畫眉通常成群活動,在樹冠層和灌木叢中靈活穿梭。它們主要以昆蟲為食,但也會食用一些漿果和花蜜。冠羽畫眉的社群行為非常有趣,它們常常一起梳理羽毛,甚至會輪流站崗警戒。
在水域環境中,台灣常見的留鳥包括小白鷺(Egretta garzetta)和翠鳥(Alcedo atthis)。小白鷺是一種優雅的水鳥,全身潔白,常在沼澤、水田和河邊覓食。它們長長的脖子和尖銳的喙特別適合捕捉魚類和其他水生生物。小白鷺雖然是留鳥,但在不同季節可能會在台灣島內進行短距離遷移。
翠鳥則是一種色彩豔麗的小型水鳥,以其亮麗的藍色上體和橙色下體而聞名。這種鳥類常在河流、溪流和魚塘邊出沒,它們會在水面上方停留,然後俯衝入水捕魚。翠鳥通常單獨或成對活動,在河岸的泥壁上挖洞築巢。
在夜行性鳥類中,領角鴞(Otus lettia)是台灣常見的留鳥之一。這種小型貓頭鷹以其特徵性的「耳朵」(實際上是羽毛)而得名。領角鴞主要生活在低海拔至中海拔的森林和農村地區,以各種昆蟲和小型脊椎動物為食。雖然它們是夜行性動物,但有時在黃昏或清晨也能看到它們的活動。
台灣的留鳥不僅豐富了當地的生物多樣性,也在生態系統中扮演著重要角色。例如,許多鳥類通過捕食昆蟲來控制害蟲數量,有些則通過傳播種子來促進植物的繁衍。同時,這些留鳥也是重要的環境指標,它們的數量和健康狀況能夠反映出棲息地的生態狀況。
然而,隨著人類活動的擴張和環境的變化,許多留鳥也面臨著挑戰。棲息地破壞、環境污染、氣候變化等因素都可能對留鳥產生負面影響。因此,保護這些鳥類及其棲息地變得尤為重要。
台灣政府和民間組織都在積極採取措施保護這些留鳥。例如,設立自然保護區和國家公園,保護重要的鳥類棲息地。同時,也開展了許多研究和監測項目,以更好地瞭解這些鳥類的生態習性和種群動態。
此外,環境教育和公眾參與也是保護留鳥的重要方面。通過舉辦觀鳥活動、製作教育材料等方式,讓更多人瞭解並欣賞這些美麗的生物,從而增強公眾的保護意識。
台灣豐富的留鳥資源不僅是寶貴的自然遺產,也為當地居民和遊客提供了欣賞自然之美的機會。無論是在繁華的城市公園,還是在寧靜的山區森林,這些留鳥都為台灣的自然環境增添了生機和活力。通過持續的保護和研究,我們希望這些美麗的生物能夠繼續在台灣的土地上繁衍生息,為子孫後代留下豐富的自然遺產。
3.10 台灣的森林鳥類
台灣是一個生物多樣性豐富的島嶼,其中森林生態系統尤為重要,孕育了眾多鳥類。台灣的森林從海平面延伸到高山,包括常綠闊葉林、落葉闊葉林、混合林和針葉林等多種類型,為不同的鳥類提供了多樣化的棲息環境。讓我們深入探討台灣常見的森林鳥類,瞭解它們的特徵、生態習性以及在生態系統中的角色。
在低海拔的常綠闊葉林中,五色鳥(Megalaima nuchalis)是一種常見且引人注目的鳥類。這種鳥以其鮮艷的羽毛而聞名,頭部和頸部呈現紅、黃、藍、綠、黑五種顏色,因此得名。五色鳥主要以果實為食,尤其喜愛無花果。牠們通常在樹冠層活動,用強壯的喙在樹幹上鑿洞築巢。五色鳥的叫聲低沉有力,在繁殖季節特別常聽到。這種鳥類在生態系統中扮演著重要的種子傳播者角色,有助於森林的更新和擴展。
另一種在低海拔森林常見的鳥類是台灣鵯(Pycnonotus taivanus)。這是台灣的特有種,主要分佈在東部和南部的低海拔森林。台灣鵯體型中等,羽毛以褐色為主,頭部有明顯的冠羽。牠們通常成小群活動,以各種果實和昆蟲為食。台灣鵯的叫聲悅耳,常在清晨和黃昏時分歌唱,為森林增添了生機。
在中海拔的混合林中,台灣藍鵲(Urocissa caerulea)是最引人注目的鳥類之一。這種美麗的鳥不僅是台灣的特有種,也是台灣的國鳥。台灣藍鵲體型較大,羽毛以亮麗的藍色為主,長長的尾巴末端有白色斑點。牠們通常成群活動,主要分佈在海拔300到1,200米的森林中。台灣藍鵲聰明機智,食性廣泛,從果實到小型脊椎動物都在其食譜之中。這種鳥類在生態系統中扮演著重要的角色,既是種子傳播者,也是小型動物的捕食者,有助於維持森林生態的平衡。
大彎嘴(Pomatorhinus erythrocnemis)是另一種在中海拔森林常見的鳥類。這種鳥是台灣的特有亞種,以其獨特的下彎長喙為特徵。大彎嘴主要生活在低海拔至中海拔的森林中,常在地面或灌木叢中覓食。牠們通常成對或小群活動,以各種昆蟲和植物種子為食。大彎嘴的叫聲清亮悅耳,是森林中常聽到的聲音之一。這種鳥類在控制森林昆蟲數量方面發揮著重要作用。
在較高海拔的森林中,冠羽畫眉(Yuhina brunneiceps)是一種常見的留鳥。這種小型鳥類是台灣的特有種,以其特徵性的直立冠羽而聞名。冠羽畫眉通常成群活動,在樹冠層和灌木叢中靈活穿梭。牠們主要以昆蟲為食,但也會食用一些漿果和花蜜。冠羽畫眉的社群行為非常有趣,牠們常常一起梳理羽毛,甚至會輪流站崗警戒。這種鳥類在高海拔森林生態系統中扮演著重要的授粉者和昆蟲控制者的角色。
台灣噪眉(Alcippe morrisonia)是另一種在中高海拔森林常見的鳥類。這種小型鳥類通常在樹冠下層和灌木叢中活動,以昆蟲和小型果實為食。台灣噪眉的羽毛以褐色為主,頭部有明顯的眉斑。牠們常常成小群活動,在樹枝間快速移動尋找食物。台灣噪眉的叫聲悅耳,是森林中常聽到的背景音樂之一。
在針葉林和高山森林中,青背山雀(Parus monticolus)是一種常見的留鳥。這種小型鳥類以其鮮艷的黃色腹部和藍綠色的背部而引人注目。青背山雀通常在樹冠層活動,以各種昆蟲和種子為食。牠們具有將食物儲存起來以備不時之需的習慣,這種行為在寒冷的高山環境中特別有利於牠們的生存。青背山雀在高山森林生態系統中扮演著重要的昆蟲控制者和種子傳播者的角色。
台灣山鷓鴣(Arborophila crudigularis)是一種在中高海拔森林常見的地棲性鳥類。這種鳥是台灣的特有種,主要生活在海拔1,000到2,500米的森林中。台灣山鷓鴣體型圓潤,羽毛以褐色為主,有著複雜的斑紋。牠們通常在地面活動,以各種植物種子和小型無脊椎動物為食。雖然台灣山鷓鴣較為害羞,不易被直接觀察到,但牠們獨特的叫聲是森林中常聽到的聲音之一。
在夜間活動的森林鳥類中,領角鴞(Otus lettia)是一種常見的留鳥。這種小型貓頭鷹以其特徵性的「耳朵」(實際上是羽毛)而得名。領角鴞主要生活在低海拔至中海拔的森林和農村地區,以各種昆蟲和小型脊椎動物為食。雖然牠們是夜行性動物,但有時在黃昏或清晨也能看到牠們的活動。領角鴞在控制森林小型動物和昆蟲數量方面發揮著重要作用。
台灣斑翅鶇(Turdus eunomus)是一種在中高海拔森林常見的鶇科鳥類。這種鳥體型中等,羽毛以褐色為主,翅膀上有明顯的白色斑紋。台灣斑翅鶇主要以各種昆蟲和漿果為食,在森林地面和灌木叢中覓食。牠們的叫聲悅耳動聽,特別是在繁殖季節,雄鳥會發出美妙的歌聲來吸引雌鳥。這種鳥類在森林生態系統中扮演著重要的種子傳播者角色。
在森林邊緣和次生林中,繡眼畫眉(Alcippe brunnea)是一種常見的小型鳥類。這種鳥以其特徵性的白色眼圈而得名。繡眼畫眉通常成小群活動,在灌木叢和低矮樹木間靈活穿梭。牠們主要以各種小型昆蟲和蜘蛛為食,有時也會食用一些漿果。繡眼畫眉的叫聲輕柔悅耳,是森林中常聽到的背景音樂之一。
在較為開闊的森林地區,台灣竹雞(Bambusicola sonorivox)是一種常見的地棲性留鳥。這種鳥類是台灣的特有種,主要分佈在低海拔至中海拔的森林和竹林中。台灣竹雞體型圓潤,羽毛以褐色為主,有著複雜的斑紋。牠們通常在地面活動,以各種植物種子和小型無脊椎動物為食。雖然台灣竹雞較為害羞,不易被直接觀察到,但牠們響亮的叫聲常能在森林中聽到。
這些森林鳥類不僅豐富了台灣的生物多樣性,也在生態系統中扮演著重要角色。牠們通過捕食昆蟲來控制害蟲數量,通過傳播種子來促進植物的繁衍,有些還通過授粉來幫助植物繁殖。同時,這些鳥類也是重要的環境指標,牠們的數量和健康狀況能夠反映出森林生態系統的整體狀況。
然而,隨著人類活動的擴張和氣候變化的影響,許多森林鳥類也面臨著各種威脅。棲息地破碎化、環境污染、非法捕獵等因素都可能對這些鳥類產生負面影響。因此,保護這些森林鳥類及其棲息地變得尤為重要。
台灣政府和民間組織都在積極採取措施保護這些森林鳥類。例如,設立自然保護區和國家公園,保護重要的森林棲息地。同時,也開展了許多研究和監測項目,以更好地瞭解這些鳥類的生態習性和種群動態。此外,環境教育和公眾參與也是保護森林鳥類的重要方面。通過舉辦觀鳥活動、製作教育材料等方式,讓更多人瞭解並欣賞這些美麗的生物,從而增強公眾的保護意識。
台灣豐富的森林鳥類資源不僅是寶貴的自然遺產,也為當地居民和遊客提供了欣賞自然之美的機會。無論是在低海拔的常綠闊葉林,還是在高山的針葉林,這些森林鳥類都為台灣的自然環境增添了生機和活力。通過持續的保護和研究,我們希望這些美麗的生物能夠繼續在台灣的森林中繁衍生息,為子孫後代留下豐富的自然遺產。
3.11 台灣的高山鳥類
台灣島嶼地形多變,高山林立,海拔1000公尺以上的區域佔據了相當大的面積。這些高山地區因其獨特的地理環境和氣候條件,孕育了許多特有的動植物種類,其中高山鳥類更是台灣生物多樣性的重要組成部分。這些鳥類不僅適應了高海拔的環境,還在生態系統中扮演著關鍵角色。讓我們深入探討台灣常見的高山鳥類,瞭解它們的特徵、生態習性以及在高山生態系統中的重要性。
台灣高山地區最具代表性的鳥類—帝雉(Syrmaticus mikado)。這種美麗的鳥類是台灣的特有種,也被稱為黑長尾雉。帝雉主要分佈在海拔2000至3000公尺的原始森林中,是台灣高山地區的明星物種。雄性帝雉有著長達1米的尾羽,全身羽毛呈現出金屬光澤的藍黑色,極為華麗。相比之下,雌性則較為樸素,呈褐色並帶有斑點,這種保護色有助於在孵卵期間隱藏自己。帝雉主要以各種植物的嫩芽、果實和種子為食,同時也會捕食一些小型無脊椎動物。由於棲息地的破壞和過去的過度捕獵,帝雉的數量曾經大幅下降,現在已被列為易危物種,受到嚴格保護。
另一種在台灣高山地區常見的鳥類是藪鳥(Liocichla steerii)。這種鳥也是台灣的特有種,主要分佈在海拔1000至3000公尺的山區。藪鳥體型中等,羽毛以橄欖綠色為主,翅膀和尾部有鮮艷的紅色和黃色。這種鳥類通常成對或小群活動,在灌木叢和低矮樹木間靈活穿梭。藪鳥的食性廣泛,包括各種漿果、種子和昆蟲。牠們的叫聲悅耳動聽,是台灣高山森林中常聽到的聲音之一。藪鳥在生態系統中扮演著重要的種子傳播者和昆蟲控制者的角色。
在台灣的高山針葉林中,青背山雀(Parus monticolus)是一種常見的留鳥。這種小型鳥類以其鮮艷的黃色腹部和藍綠色的背部而引人注目。青背山雀主要分佈在海拔1500至3000公尺的山區,牠們通常在樹冠層活動,以各種昆蟲和種子為食。青背山雀有一個有趣的習性,就是會將食物儲存起來以備不時之需,這種行為在寒冷的高山環境中特別有利於牠們的生存。這種鳥類在高山森林生態系統中扮演著重要的昆蟲控制者和種子傳播者的角色。
冠羽畫眉(Yuhina brunneiceps)是另一種在台灣高山地區常見的鳥類。這種小型鳥類是台灣的特有種,以其特徵性的直立冠羽而聞名。冠羽畫眉主要分佈在海拔1000至3000公尺的山區,通常成群活動,在樹冠層和灌木叢中靈活穿梭。牠們主要以昆蟲為食,但也會食用一些漿果和花蜜。冠羽畫眉的社群行為非常有趣,牠們常常一起梳理羽毛,甚至會輪流站崗警戒。這種鳥類在高山森林生態系統中扮演著重要的授粉者和昆蟲控制者的角色。
台灣噪眉(Alcippe morrisonia)也是台灣高山地區常見的鳥類之一。這種小型鳥類通常在海拔1000至2500公尺的山區活動,牠們喜歡在樹冠下層和灌木叢中穿梭,以昆蟲和小型果實為食。台灣噪眉的羽毛以褐色為主,頭部有明顯的眉斑。牠們常常成小群活動,在樹枝間快速移動尋找食物。台灣噪眉的叫聲悅耳,是高山森林中常聽到的背景音樂之一。
在台灣的高山草原和森林邊緣地帶,白尾鴝(Phoenicurus auroreus)是一種常見的鳥類。這種鳥主要在海拔1500至3000公尺的地區活動,雄鳥有著醒目的橙紅色胸部和黑色背部,而雌鳥則較為樸素,呈褐色。白尾鴝喜歡在開闊地帶活動,常常可以看到牠們站在突出的枝條或岩石上,警惕地觀察四周。牠們主要以各種昆蟲為食,有時也會吃一些小型漿果。白尾鴝在高山生態系統中扮演著重要的昆蟲控制者的角色。
台灣山鷓鴣(Arborophila crudigularis)是另一種在台灣高山地區常見的地棲性鳥類。這種鳥是台灣的特有種,主要生活在海拔1000至2500公尺的森林中。台灣山鷓鴣體型圓潤,羽毛以褐色為主,有著複雜的斑紋。牠們通常在地面活動,以各種植物種子和小型無脊椎動物為食。雖然台灣山鷓鴣較為害羞,不易被直接觀察到,但牠們獨特的叫聲是高山森林中常聽到的聲音之一。這種鳥類在高山森林生態系統中扮演著重要的種子傳播者和昆蟲控制者的角色。
在台灣的高山針葉林中,星鴉(Nucifraga caryocatactes)是一種引人注目的鳥類。這種鳥體型較大,羽毛以深褐色為主,全身佈滿白色斑點,看起來就像滿天星斗,因此得名。星鴉主要分佈在海拔2000至3500公尺的高山地區,牠們的食性主要以松樹和杉樹的種子為主,因此在生態系統中扮演著重要的種子傳播者角色。星鴉有一個有趣的習性,就是會將多餘的種子儲存起來,這種行為不僅幫助牠們度過食物匱乏的時期,也促進了森林的自然更新。
高山地區的猛禽中,熊鷹(Nisaetus nipalensis)是一種常見的大型猛禽。這種鳥主要分佈在海拔1000至3000公尺的山區,是台灣最大的林下獵食性猛禽。熊鷹有著強壯的身軀和銳利的爪子,主要以中小型哺乳動物和鳥類為食。牠們在生態系統中扮演著頂級捕食者的角色,對維持生態平衡至關重要。由於棲息地的減少和人為幹擾,熊鷹的數量在過去曾經大幅下降,現在已被列為易危物種,受到嚴格保護。
3.12 台灣 的地棲性 鳥類
台灣最常見的地棲性鳥類之一—台灣竹雞(Bambusicola sonorivox)。這種鳥是台灣的特有種,主要分佈在低海拔至中海拔的森林和竹林中。台灣竹雞體型圓潤,羽毛以褐色為主,有著複雜的斑紋,這種保護色使得牠們能夠很好地隱藏在地表植被中。台灣竹雞通常成對或小群活動,以各種植物種子、嫩芽和小型無脊椎動物為食。雖然台灣竹雞較為害羞,不易被直接觀察到,但牠們響亮的叫聲是森林中常聽到的聲音之一。這種鳥類在生態系統中扮演著重要的種子傳播者和昆蟲控制者的角色。
環頸雉(Phasianus colchicus)廣泛分佈在台灣的平原和低海拔丘陵地區,特別是在農田和草地附近。雄性環頸雉有著鮮艷的羽毛,頭部呈綠色,頸部有一圈白色環狀斑紋,身體呈銅紅色,尾部特別長。相比之下,雌性則較為樸素,呈褐色並帶有斑點,這種保護色有助於在孵卵期間隱藏自己。環頸雉主要以各種植物種子、嫩芽和昆蟲為食,在農業生態系統中扮演著重要的害蟲控制者角色。
在台灣的高山地區,台灣山鷓鴣(Arborophila crudigularis)是一種常見的地棲性鳥類。這種鳥是台灣的特有種,主要生活在海拔1000至2500公尺的森林中。台灣山鷓鴣體型圓潤,羽毛以褐色為主,有著複雜的斑紋。牠們通常在地面活動,以各種植物種子和小型無脊椎動物為食。雖然台灣山鷓鴣較為害羞,不易被直接觀察到,但牠們獨特的叫聲是高山森林中常聽到的聲音之一。這種鳥類在高山森林生態系統中扮演著重要的種子傳播者和昆蟲控制者的角色。
在台灣的沿海地區和濕地,麥雞(Turnicidae)是一種常見的小型地棲性鳥類。麥雞有著圓潤的身體和短小的尾巴,羽毛以褐色為主,有著複雜的斑紋。有趣的是,在麥雞中,雌性比雄性更鮮艷,而且在繁殖期間,雌性負責求偶和保衛領地,雄性則負責孵卵和照顧幼鳥。麥雞主要以各種種子和小型無脊椎動物為食,在濕地生態系統中扮演著重要的角色。
黑面琵鷺(Platalea minor)雖然不是嚴格意義上的地棲性鳥類,但牠們大部分時間都在淺水區域或泥灘上活動,因此也可以被視為一種特殊的地棲性鳥類。這種全球瀕危物種每年冬天會來到台灣南部的沿海濕地越冬。黑面琵鷺有著獨特的勺狀喙,用來在淺水中掃食小魚、蝦和其他水生生物。牠們的到來不僅豐富了台灣的生物多樣性,也成為了濕地生態健康的重要指標。
在台灣的開闊草地和農田地區,小環頸鴴(Charadrius dubius)是一種常見的地棲性鳥類。這種鳥體型小巧,羽毛以褐色為主,胸部有一圈黑色環狀斑紋。小環頸鴴通常在地面快速奔跑,停下來時會突然靜止,這種行為有助於牠們躲避天敵。牠們主要以各種小型無脊椎動物為食,在控制害蟲方面發揮著重要作用。
田鷸(Rostratula benghalensis)是另一種在台灣濕地常見的地棲性鳥類。這種鳥有著獨特的外表,雌鳥比雄鳥更鮮艷,有著褐色的羽毛和醒目的白色環眼。田鷸主要在泥濘的地面和淺水區域覓食,以各種水生無脊椎動物為食。牠們長而彎曲的喙特別適合在軟泥中尋找食物。田鷸在濕地生態系統中扮演著重要的角色,其存在往往意味著濕地生態的健康。
在台灣的森林地區,深山竹雞(Arborophila crudigularis)是一種常見的地棲性鳥類。這種鳥是台灣的特有種,主要分佈在中高海拔的森林中。深山竹雞體型略大於台灣竹雞,羽毛以褐色為主,有著複雜的斑紋。牠們通常在森林地面活動,以各種植物種子、漿果和小型無脊椎動物為食。深山竹雞的叫聲低沉有力,是山區森林中常聽到的聲音之一。
在台灣的沿海地區,東方環頸鴴(Charadrius alexandrinus)是一種常見的地棲性鳥類。這種鳥體型小巧,羽毛以灰褐色為主,頸部有一圈不完整的黑色環狀斑紋。東方環頸鴴主要在沙灘和泥灘上活動,以各種小型無脊椎動物為食。牠們有一個有趣的習性,就是會使用「受傷鳥」戰術來吸引天敵遠離巢穴,保護幼鳥。
在台灣的農田和草地地區,白鶺鴒(Motacilla alba)是一種常見的地棲性鳥類。這種鳥體型修長,羽毛以黑白兩色為主,尾巴特別長,常常上下擺動。白鶺鴒主要在地面活動,以各種小型昆蟲為食。牠們經常可以看到在牛隻附近活動,捕食被牛隻驚起的昆蟲。白鶺鴒在農業生態系統中扮演著重要的害蟲控制者角色。
3.13 台灣的都市棲息鳥類
台灣的都市化進程雖然改變了許多自然棲息地,但同時也創造了新的生態環境,吸引了一些適應性強的鳥類在此定居。這些都市棲息鳥類不僅豐富了城市的生態系統,也為都市居民帶來了親近自然的機會。讓我們深入探討台灣常見的都市棲息鳥類,瞭解它們的特徵、生活習性,以及在都市生態中扮演的角色。
麻雀(Passer montanus)無疑是台灣都市中最常見的鳥類之一。這種小型鳥類已經很好地適應了人類環境,幾乎在每個城市的角落都能看到它們的身影。麻雀體型小巧,羽毛以褐色為主,頭頂和後頸呈栗色,臉頰有明顯的黑斑。它們通常成群活動,在建築物的縫隙、屋簷下或樹洞中築巢。麻雀的食性廣泛,既吃各種植物種子,也不排斥人類食物的殘渣,這種適應性使得它們能夠在都市環境中茁壯成長。麻雀的存在不僅為城市增添了生氣,也在一定程度上幫助控制了昆蟲數量。
白頭翁(Pycnonotus sinensis)是另一種在台灣都市中常見的鳥類。這種中型鳥類以其特徵性的白色頭部和黑色冠羽而聞名。白頭翁的叫聲悅耳動聽,常在清晨和黃昏時分歌唱,為都市的喧囂帶來一絲自然的和諧。它們主要以果實和昆蟲為食,因此在城市公園、果園和住宅區的花園中經常可以看到它們的身影。白頭翁適應性強,在城市和鄉村地區都能生存良好,是台灣最常見的鳥類之一。
樹鵲(Dendrocitta formosae)是一種在台灣都市中引人注目的鳥類。這種鳥體型較大,羽毛以黑白兩色為主,長長的尾巴特別醒目。樹鵲聰明機智,食性廣泛,從果實、昆蟲到小型脊椎動物都是它們的食物。在都市環境中,樹鵲常常可以在公園、校園或有大樹的住宅區看到。它們的叫聲響亮,有時會模仿其他鳥類或環境聲音,為都市增添了幾分野趣。
綠繡眼(Zosterops japonicus)是一種小型鳥類,在台灣的都市環境中也很常見。這種鳥以其鮮艷的黃綠色羽毛和明顯的白色眼圈為特徵。綠繡眼通常成小群活動,它們敏捷靈活,常在樹枝間跳躍覓食。除了昆蟲外,綠繡眼還喜歡吸食花蜜,因此在開花的季節,我們常能看到它們在城市公園或住宅區的花園中忙碌的身影。綠繡眼的存在不僅為都市增添了色彩,也在植物授粉方面發揮了重要作用。
大卷尾(Dicrurus macrocercus)是一種在台灣都市中常見的黑色鳥類。這種鳥體型中等,全身羽毛呈亮麗的黑色,最引人注目的是它那叉狀的長尾。大卷尾常常可以看到停在電線或樹枝上,警惕地觀察四周。它們主要以飛蟲為食,常常在空中靈活地追逐獵物。在都市環境中,大卷尾扮演著重要的昆蟲控制者角色,幫助維持生態平衡。
家燕(Hirundo rustica)和赤腰燕(Cecropis daurica)是台灣都市中常見的燕子。這兩種鳥類通常在春季抵達台灣繁殖,秋季再南返越冬。它們優雅的飛行姿態和叉狀的尾部是其顯著特徵。燕子常在建築物下築巢,利用泥土和唾液建造巢穴。它們主要以飛蟲為食,常常可以看到它們在城市上空靈活地飛翔捕蟲。燕子的存在不僅為都市增添了生機,也幫助控制了蚊蠅等害蟲的數量。
斑鳩(Spilopelia chinensis)是另一種在台灣都市中常見的鳥類。這種鳥體型中等,羽毛以褐色為主,頸部有明顯的黑白條紋。斑鳩通常成對或小群活動,常在地面覓食,以各種植物種子為主。在都市環境中,斑鳩常常可以在公園、校園或住宅區的草地上看到。它們柔和的叫聲為都市增添了一份寧靜的氛圍。
喜鵲(Pica pica)雖然不如其他鳥類那麼普遍,但在某些都市地區也是常見的鳥類。這種黑白相間的大型鳥類以其長尾和金屬光澤的羽毛而引人注目。喜鵲聰明機智,食性廣泛,從果實、昆蟲到小型脊椎動物都是它們的食物。在都市環境中,喜鵲常常可以在有大樹的公園或郊區看到。它們的存在為都市增添了幾分野趣。
八哥(Acridotheres cristatellus)是一種在台灣都市中常見的鳥類,儘管它是外來種。這種鳥體型中等,羽毛以黑色為主,嘴和腳呈黃色。八哥聰明活潑,學舌能力強,食性廣泛,能夠很好地適應都市環境。它們常常成群出現在公園、廣場或有大樹的街道上。雖然八哥的入侵對本地鳥類造成了一定的競爭壓力,但它們也成為了都市景觀中不可或缺的一部分。
夜鷺(Nycticorax nycticorax)是一種在台灣都市中常見的夜行性鳥類。這種鳥體型中等,羽毛以灰色和白色為主,頭頂和背部呈黑色。夜鷺通常在黃昏時分開始活動,常常可以看到它們停在都市中的大樹或建築物上。它們主要以魚類和其他水生生物為食,因此在都市中的河流、湖泊或公園的水池附近常能看到它們的身影。夜鷺的存在豐富了都市的夜間生態。
這些都市棲息鳥類不僅豐富了台灣城市的生物多樣性,也在都市生態系統中扮演著重要角色。它們通過捕食昆蟲來控制害蟲數量,通過傳播種子來促進植物的繁衍,有些還通過授粉來幫助植物繁殖。同時,這些鳥類也是重要的環境指標,它們的數量和健康狀況能夠反映出都市生態系統的整體狀況。
然而,都市棲息鳥類也面臨著諸多挑戰。城市化進程導致的棲息地破碎化和消失是一大威脅。此外,建築物玻璃的反光造成的鳥類撞擊、光污染對夜行性鳥類的幹擾、以及城市中流浪貓狗的捕食等,都是都市鳥類面臨的問題。空氣污染和噪音污染也可能對鳥類的健康和繁殖造成負面影響。
為了保護這些都市棲息鳥類,台灣的政府和民間組織採取了多項措施。例如,在城市規劃中保留更多的綠地和水域,為鳥類提供棲息和覓食的場所。鼓勵市民在陽台或庭院種植適合鳥類的植物,為鳥類創造微型棲息地。同時,也開展了許多研究和監測項目,以更好地瞭解都市鳥類的生態習性和種群動態。
環境教育和公眾參與也是保護都市鳥類的重要方面。通過舉辦都市觀鳥活動、製作教育材料等方式,讓更多市民瞭解並欣賞身邊的鳥類,從而增強公眾的保護意識。許多學校和社區組織也開展了都市鳥類教育項目,讓孩子們從小就瞭解都市生態的重要性。
都市棲息鳥類的存在不僅豐富了城市的生態系統,也為都市居民帶來了親近自然的機會。它們的鳴叫聲打破了城市的喧囂,它們的飛翔為灰色的水泥叢林增添了生機。通過保護這些都市鳥類,我們不僅是在維護生物多樣性,也是在為自己創造一個更宜居、更有生命力的城市環境。都市和自然並非對立,而是可以和諧共存的。都市棲息鳥類的繁榮正是這種和諧共存的最好證明。
3.14 台灣的河口濱海水鳥
台灣四面環海,擁有豐富的河口和濱海生態系統,為眾多水鳥提供了理想的棲息和覓食環境。這些區域不僅是許多留鳥的家園,也是候鳥遷徙路線上的重要中繼站。讓我們深入探討台灣常見的河口和濱海水鳥,包括游禽和涉禽,瞭解它們的特徵、生態習性以及在這些濕地生態系統中的重要性。
常見的游禽-小水鴨(Anas crecca)是台灣冬季常見的候鳥之一。這種鴨子體型較小,雄鳥頭部有著醒目的綠色羽毛,身體呈灰褐色,翅膀上有一塊綠色的鏡羽。小水鴨通常成群出現在河口、濕地和沿海淺水區域,主要以水生植物、小型無脊椎動物和種子為食。每年秋季,大批小水鴨會從北方飛來台灣過冬,為寒冷的季節增添了生機。
綠頭鴨(Anas platyrhynchos)是另一種在台灣河口和濱海地區常見的游禽。雄鳥有著標誌性的綠色頭部和黃色喙,身體呈灰褐色,尾部有捲曲的黑色羽毛。綠頭鴨適應性強,能在各種水域環境中生存,包括河口、濕地、湖泊和沿海地區。它們的食性廣泛,從水生植物到小型動物都在其食譜之中。綠頭鴨不僅是重要的生態系統成員,也是常見的狩獵對象,因此其數量的變化受到密切關注。
琵嘴鴨(Spatula cpeata)是一種特別的鴨子,以其寬大的鏟形喙而聞名。這種鴨子在台灣主要是冬候鳥,常見於河口和沿海濕地。雄鳥在繁殖羽時有著綠色的頭部、白色的胸部和栗色的腹部,非常醒目。琵嘴鴨的特殊喙部構造使它們能夠有效地在水中過濾食物,主要捕食浮游生物和小型水生無脊椎動物。
在涉禽中,高蹺鴴(Himantopus himantopus)是一種常見且引人注目的鳥類。這種鳥有著極長的紅色腿部,黑白分明的羽毛,站立時顯得優雅而高挑。高蹺鴴喜歡在淺水區域覓食,主要捕食水生昆蟲、小魚和甲殼類動物。它們常見於河口、魚塭和沿海濕地,其獨特的外形使它們成為了濕地生態系統中的標誌性物種之一。
反嘴鴴(Recurvirostra avosetta)是另一種特別的涉禽,以其向上彎曲的長喙而聞名。這種鳥羽毛呈黑白相間,有著修長的淺藍色腿部。反嘴鴴主要在淺水區域和泥灘上覓食,利用其獨特的喙部在水中左右擺動,濾食小型水生生物。它們常見於河口和沿海濕地,是台灣重要的冬候鳥之一。
東方環頸鴴(Charadrius alexandrinus)是一種小型涉禽,在台灣的河口和海岸地區常年可見。這種鳥體型嬌小,羽毛以灰褐色為主,頸部有一圈不完整的黑色環狀斑紋。東方環頸鴴主要在沙灘和泥灘上活動,以各種小型無脊椎動物為食。它們有一個有趣的習性,就是會使用「受傷鳥」戰術來吸引天敵遠離巢穴,保護幼鳥。
黑面琵鷺(Platalea minor)是台灣河口和濱海地區最知名的涉禽之一。這種全球瀕危物種每年冬天會來到台灣南部的沿海濕地越冬。黑面琵鷺有著獨特的勺狀喙,用來在淺水中掃食小魚、蝦和其他水生生物。牠們的到來不僅豐富了台灣的生物多樣性,也成為了濕地生態健康的重要指標。台灣在黑面琵鷺的保育中扮演著關鍵角色,每年到訪的數量約占全球總數的一半左右。
大白鷺(Ardea alba)和中白鷺(Egretta intermedia)是台灣河口和濱海地區常見的大型涉禽。這些鷺鳥以其純白的羽毛和修長的頸部聞名,常在淺水區域覓食,捕捉魚類和其他水生生物。雖然這些鷺鳥在全球範圍內分佈廣泛,但它們在台灣的大量聚集仍然是濕地生態系統的重要組成部分。
鷸科鳥類如黑腹濱鷸(Calidris alpina)和磯鷸(Actitis hypoleucos)也是台灣河口和濱海地區常見的涉禽。這些鳥類通常體型較小,有著長而尖的喙,適合在泥灘和沙灘上覓食。牠們主要以各種無脊椎動物為食,在潮間帶的生態系統中扮演著重要角色。每年秋冬季節,大量的鷸鴴類會途經或在台灣停留,形成了壯觀的遷徙景象。
鸕鶿(Phalacrocorax carbo)雖然不是涉禽,但作為一種特殊的游禽,在台灣的河口和濱海地區也很常見。這種大型水鳥全身羽毛呈黑色,有著長而彎曲的喙,善於潛水捕魚。鸕鶿常常可以看到停在岩石或樁柱上,張開翅膀晾乾羽毛的樣子。牠們在水域生態系統中扮演著重要的頂級消費者角色。
赤頸鴨(Mareca penelope)是另一種在台灣河口和濱海地區常見的游禽。這種鴨子是重要的冬候鳥,通常在秋季抵達台灣。雄鳥有著醒目的栗色頭部和淺灰色的身體,而雌鳥則較為樸素,呈褐色。赤頸鴨主要以水生植物為食,但也會捕食一些小型水生動物。牠們常在開闊的水域活動,為冬季的濕地增添了生機。
在鷗科鳥類中,黑尾鷗(Larus crassirostris)和紅嘴鷗(Chroicocephalus ridibundus)是台灣沿海地區常見的種類。這些海鳥以其優雅的飛行姿態和響亮的叫聲而聞名。牠們主要以魚類和其他海洋生物為食,常在海岸線附近活動。鷗類在海洋生態系統中扮演著重要的角色,其數量和分佈的變化常被用來指示海洋環境的健康狀況。
3.15 台灣的內陸溪流水鳥
台灣地形多樣,內陸溪流與湖塘遍佈全島,為眾多鳥類提供了理想的棲息、覓食和築巢環境。這些水域及其周邊地區不僅是許多留鳥的家園,也吸引了不少候鳥前來停留。讓我們深入探討台灣常見的內陸溪流湖塘周邊鳥類,瞭解它們的特徵、生態習性以及在這些淡水生態系統中的重要性。
翠鳥(Alcedo atthis),這種色彩豔麗的小型鳥類是台灣內陸水域最引人注目的居民之一。翠鳥全身羽毛呈亮麗的藍色和橙色,有著尖銳的長喙,非常適合捕魚。它們常常可以看到停在溪流或湖泊邊的枝條上,專注地盯著水面,一旦發現獵物就會迅速俯衝入水捕捉。翠鳥通常在河岸的泥壁上挖掘隧道狀的巢穴,深入可達一米左右。這種獨特的築巢方式為它們的幼鳥提供了安全的生長環境。翠鳥不僅是一種美麗的鳥類,也是淡水生態系統健康的重要指標。
小白鷺(Egretta garzetta)是另一種在台灣內陸水域常見的鳥類。這種優雅的白色水鳥體型中等,有著細長的頸部和尖銳的黑色喙。小白鷺常在溪流、湖泊和魚塭等淺水區域覓食,主要捕食小魚、蝦和水生昆蟲。在繁殖季節,小白鷺會聚集在水邊的樹林中築巢,形成鷺鷥林。這種集體築巢的行為不僅能夠提高育雛的成功率,也為生態系統帶來了豐富的養分輸入。小白鷺適應性強,能在各種水域環境中生存,是台灣淡水生態系統中的重要成員。
河烏(Cinclus pallasii)是一種特別的鳥類,專門適應了湍急的山溪環境。這種鳥全身羽毛呈深褐色,胸部有一塊白斑。河烏最引人注目的特點是它們能夠在急流中游泳和潛水,甚至能夠逆流而上。它們主要以水生昆蟲和小魚為食,常常可以看到它們站在溪中的岩石上,不斷下潛覓食的樣子。河烏通常在瀑布或急流附近的岩壁縫隙中築巢,巢穴常常被水花濺濕。這種鳥類的存在往往意味著溪流水質清澈,生態環境良好。
在台灣的湖泊和魚塭周圍,常常可以看到鸕鶿(Phalacrocorax carbo)的身影。這種大型水鳥全身羽毛呈黑色,有著長而彎曲的喙,善於潛水捕魚。鸕鶿常常可以看到停在岩石或樁柱上,張開翅膀晾乾羽毛的樣子。它們通常在水邊的大樹上築巢,有時會形成大型的群體繁殖地。鸕鶿在水域生態系統中扮演著重要的頂級消費者角色,其數量的變化能夠反映出魚類資源的豐富程度。
黃頭鷺(Bubulcus ibis)是一種適應性極強的鳥類,不僅在沿海地區常見,在內陸的湖泊、溪流和農田周圍也經常可以看到它們的身影。這種鷺鳥體型較小,全身羽毛呈白色,在繁殖季節頭部、胸部和背部會呈現金黃色。黃頭鷺常常可以看到跟隨在牛群周圍覓食,捕食被牛隻驚起的昆蟲。它們也會在淺水區域捕食小魚和兩棲類動物。黃頭鷺通常在水邊的樹林中集體築巢,有時會與其他鷺鳥物種共用一個鷺鷥林。
山紅頭(Cyornis vivida)是一種在台灣中低海拔溪流周邊常見的鳥類。這種小型鳥類雄鳥有著鮮豔的橙紅色頭部和藍色的背部,非常引人注目。山紅頭喜歡在溪流附近的林下層活動,主要以昆蟲為食。它們會利用樹洞或岩壁縫隙築巢,有時也會使用人工鳥巢箱。山紅頭悅耳的鳴叫聲是山溪環境中常聽到的聲音之一,為寧靜的溪穀增添了生機。
台灣擁有多個位於高海拔的湖泊,如翠峰湖和鴛鴦湖等,這些地方孕育了一些特殊的鳥類。其中,黃足鷸(Tringa incana)是一種在高山湖泊邊常見的鳥類。這種中型涉禽羽毛呈灰褐色,有著細長的喙和腿。金色日鷸通常在湖岸的淺水區域和泥灘上覓食,捕食各種水生無脊椎動物。雖然它們主要是過境鳥,但在適合的環境中也會有少量個體留下來繁殖。
灰鶺鴒(Motacilla cinerea)是另一種在台灣溪流邊常見的鳥類。這種小型鳥類有著優雅的身形,長長的尾巴不斷上下擺動,是它的標誌性動作。灰鶺鴒喜歡在溪流邊的石頭或淺水區活動,主要捕食水生和陸生昆蟲。它們常在溪流附近的岩壁縫隙或橋樑下方築巢。灰鶺鴒的存在通常意味著溪流環境清潔,生態系統健康。
在一些較大的湖泊和水庫周圍,常常可以看到魚鷹(Pandion haliaetus)的身影。這種大型猛禽專門捕食魚類,有著有力的爪子和彎曲的喙。魚鷹會在水面上盤旋,一旦發現獵物就會俯衝入水捕魚。它們通常在水邊的大樹或人工構築物上築巢,巢穴常年使用並不斷擴建,可以變得非常巨大。魚鷹的存在不僅豐富了湖泊生態系統,也為我們提供了瞭解水域生態健康狀況的重要線索。
台灣水雉(Jacana jacana)是一種特殊的水鳥,主要分佈在台灣中南部的埤塘和沼澤地區。這種鳥有著特別長的腳趾,能夠在浮葉植物上行走如履平地。台灣水韭以水生昆蟲和小型軟體動物為食,在覓食時常常可以看到它們在荷葉或布袋蓮上優雅行走的樣子。有趣的是,在繁殖季節,雌性水韭會主動求偶,而雄性負責孵卵和照顧幼鳥。
池鷺(Ardeola bacchus)是另一種在台灣內陸水域常見的鷺鳥。這種中小型鷺鳥在非繁殖季節時全身呈褐色,但在繁殖季節頭部和背部會變成漂亮的栗色。池鷺常在淺水區域或草叢中靜止不動,伺機捕捉經過的獵物。它們的食物包括魚類、兩棲類和大型昆蟲。池鷺通常在水邊的樹林中築巢,有時會加入混種鷺鷥林。
3.16 台灣的猛禽
台灣的猛禽是島嶼生態系統中的重要成員,它們作為食物鏈的頂級捕食者,在維持生態平衡中扮演著關鍵角色。台灣的地理位置和多樣化的地形為各種猛禽提供了豐富的棲息環境,從海岸到高山,都有不同種類的猛禽分佈。讓我們深入瞭解台灣常見的猛禽,探討它們的特徵、生態習性以及在台灣生態系統中的重要性。
首先,我們來談談台灣最大的猛禽之一—熊鷹(Nisaetus nipalensis)。這種強壯的鷹類主要分佈在台灣中低海拔的森林地區,體長可達70釐米,翼展可達1.5米。熊鷹有著深褐色的羽毛,頭部有明顯的冠羽,強而有力的爪子非常適合捕捉獵物。它們主要以中小型哺乳動物和鳥類為食,如鼬獾、松鼠和山鷓鴣等。熊鷹通常在大型喬木上築巢,每年只育養一隻幼鳥。由於棲息地減少和人為幹擾,熊鷹的數量在過去曾經大幅下降,現已被列為易危物種,受到嚴格保護。
鳳頭蒼鷹(Accipiter trivirgatus)是另一種在台灣常見的林下猛禽。這種中型猛禽體型略小於熊鷹,但同樣擁有敏捷的飛行能力和銳利的視力。鳳頭蒼鷹的頭部有明顯的羽冠,背部呈灰褐色,腹部有褐色橫紋。它們主要在森林中獵捕小型鳥類和松鼠等小型哺乳動物。鳳頭蒼鷹通常選擇在高大的樹木上築巢,巢穴常年使用並不斷修補。這種猛禽適應性較強,在城市郊區的綠地中也能生存。
大冠鷲(Spilornis cheela)是台灣特有亞種,也是常見的大型猛禽之一。這種鷲鳥全身羽毛呈深褐色,腹部和翅膀下方有明顯的白色斑點,頭部有一個顯著的羽冠。大冠鷲主要分佈在低海拔到中海拔的森林地區,它們的主要食物是蛇類,因此也被稱為"蛇鷹"。大冠鷲常在空中盤旋,利用銳利的視力搜尋地面的獵物。它們通常在大樹上築巢,每年只育養一隻幼鳥。
黑鳶(Milvus migrans)是台灣常見的中型猛禽,全身羽毛呈暗褐色,尾部略呈叉形。這種猛禽適應性極強,不僅在鄉村地區常見,在城市中也經常可以看到它們的身影。黑鳶的食性非常廣泛,從小型哺乳動物、鳥類到魚類,甚至包括腐肉和人類丟棄的食物殘渣。它們常在空中盤旋,利用敏銳的視力尋找食物。黑鳶通常在大樹或高聳的人工建築物上築巢。
遊隼(Falco peregrinus)是世界上速度最快的鳥類,在台灣主要作為冬候鳥出現。這種中型猛禽以其俯衝捕食的技巧而聞名,速度可達每小時320公里。遊隼有著深灰色的背部和白色帶有黑色橫紋的腹部,臉部有明顯的「鬍鬚」狀黑色斑紋。它們主要捕食中小型鳥類,常在海岸、濕地和開闊地區活動。近年來,遊隼已經開始在台灣的一些城市中築巢繁殖,利用高樓作為棲息和獵食的場所。
赤腹鷹(Accipiter soloensis)是台灣重要的過境猛禽。每年春秋兩季,大量的赤腹鷹會途經台灣,特別是在恆春半島,形成壯觀的遷徙景象。這種小型猛禽雄鳥上體呈灰色,腹部呈淡紅色,雌鳥則較為樸素。赤腹鷹主要以昆蟲為食,尤其喜歡蝗蟲,因此在農業生態系統中扮演著重要的害蟲控制者角色。
魚鷹(Pandion haliaetus)是一種特化的漁獵專家,在台灣的海岸、河口和大型湖泊周圍常見。這種中型猛禽有著白色的頭部和腹部,背部呈深褐色,眼睛周圍有一圈黑色羽毛。魚鷹有著特殊的腳爪結構,非常適合抓握滑溜的魚類。它們會在水面上盤旋,一旦發現獵物就會俯衝入水捕魚。魚鷹通常在水邊的大樹或人工構築物上築巢,巢穴常年使用並不斷擴建,可以變得非常巨大。
台灣最小的猛禽是紅腳隼(Falco vespertinus),這種小型隼類主要是過境鳥,在春秋兩季途經台灣。雄鳥全身呈暗灰色,腿部和眼圈呈鮮紅色,非常醒目。紅腳隼主要以昆蟲為食,常在黃昏時分群體獵食飛蟲,飛行姿態優美靈活。
除了這些日行性猛禽,台灣還有幾種常見的貓頭鷹。領角鴞(Otus lettia)是其中最常見的一種,這種小型貓頭鷹以其特徵性的「耳朵」(實際上是羽毛)而得名。領角鴞主要生活在低海拔至中海拔的森林和農村地區,以各種昆蟲和小型脊椎動物為食。雖然它們是夜行性動物,但有時在黃昏或清晨也能看到它們的活動。
3.17 台灣的鳴禽
台灣的鳴禽,也就是雀形目鳥類,構成了島上鳥類多樣性的重要部分。這些鳥類不僅數量眾多,種類豐富,還以其悅耳的鳴叫聲和多彩的羽毛為台灣的自然環境增添了無限生機。讓我們深入探討台灣常見的鳴禽,瞭解它們的特徵、生態習性以及在生態系統中的角色。
台灣最常見的鳴禽之一—麻雀(Passer montanus)。這種小型鳥類已經很好地適應了人類環境,幾乎在台灣的每個角落都能看到它們的身影。麻雀體型小巧,羽毛以褐色為主,頭頂和後頸呈栗色,臉頰有明顯的黑斑。它們通常成群活動,在建築物的縫隙、屋簷下或樹洞中築巢。麻雀的食性廣泛,既吃各種植物種子,也不排斥人類食物的殘渣,這種適應性使得它們能夠在都市環境中茁壯成長。
白頭翁(Pycnonotus sinensis)是另一種在台灣非常普遍的鳴禽。這種中型鳥類以其特徵性的白色頭部和黑色冠羽而聞名。白頭翁的叫聲悅耳動聽,常在清晨和黃昏時分歌唱,為都市和鄉村地區增添了一份自然的和諧。它們主要以果實和昆蟲為食,因此在果園和公園中經常可以看到它們的身影。白頭翁適應性強,在城市和鄉村地區都能生存良好,是台灣最常見的鳥類之一。
綠繡眼(Zosterops japonicus)是一種小型鳴禽,在台灣的各種環境中都很常見。這種鳥以其鮮艷的黃綠色羽毛和明顯的白色眼圈為特徵。綠繡眼通常成小群活動,它們敏捷靈活,常在樹枝間跳躍覓食。除了昆蟲外,綠繡眼還喜歡吸食花蜜,因此在開花的季節,我們常能看到它們在花叢中忙碌的身影。綠繡眼的存在不僅為環境增添了色彩,也在植物授粉方面發揮了重要作用。
烏頭翁(Megacapitalism canorus),是台灣特有種,主要分佈在中低海拔山區。這種鳥全身羽毛呈暗褐色,頭部略帶黑色,有著悅耳的鳴叫聲。小卡腦格主要以昆蟲和漿果為食,在森林邊緣和次生林中常見。它們通常在灌木叢中築巢,對棲息地的選擇較為靈活。
台灣畫眉(Garrulax taewanus)是另一種台灣特有種鳴禽,主要分佈在中低海拔的森林地區。這種鳥體型中等,羽毛以褐色為主,頭部和胸部有明顯的白色斑紋。台灣畫眉有著優美的歌聲,常常被稱為「百靈鳥」。它們主要以昆蟲和小型無脊椎動物為食,在林下層活動。由於棲息地的減少,台灣畫眉的數量已經大幅下降,現已被列為瀕危物種。
五色鳥(Megalaima nuchalis)雖然不是典型的鳴禽,但作為雀形目鳥類的一員,也值得一提。這種鳥以其鮮艷的羽毛而聞名,頭部和頸部呈現紅、黃、藍、綠、黑五種顏色。五色鳥主要以果實為食,特別喜愛無花果。它們通常在樹冠層活動,用強壯的喙在樹幹上鑿洞築巢。五色鳥的叫聲低沉有力,在繁殖季節特別常聽到。
樹鵲(Dendrocitta formosae)是台灣常見的大型鳴禽。這種鳥羽毛以黑白兩色為主,長長的尾巴特別醒目。樹鵲聰明機智,食性廣泛,從果實、昆蟲到小型脊椎動物都是它們的食物。在都市環境中,樹鵲常常可以在公園、校園或有大樹的住宅區看到。它們的叫聲響亮,有時會模仿其他鳥類或環境聲音。
繡眼畫眉(Alcippe morrisonia)是台灣中低海拔森林中常見的小型鳴禽。這種鳥羽毛以褐色為主,眼睛周圍有一圈白色羽毛,因而得名。繡眼畫眉通常成小群活動,在灌木叢和低矮樹木間靈活穿梭。它們主要以昆蟲為食,也會吃一些小型果實。繡眼畫眉的叫聲清脆悅耳,是森林中常聽到的聲音之一。
紅嘴黑鵯(Hypsipetes leucocephalus)是台灣中低海拔地區常見的中型鳴禽。這種鳥全身羽毛呈深灰色,頭部略呈黑色,最顯著的特徵是鮮紅色的喙。紅嘴黑鵯主要以果實和昆蟲為食,常在森林邊緣和果園活動。它們的叫聲響亮,常常成群出現,為環境增添了不少生氣。
小彎嘴(Pomatorhinus musicus)是台灣特有種,主要分佈在中低海拔的森林中。這種鳥以其獨特的下彎長喙為特徵,羽毛以褐色為主,腹部呈白色。小彎嘴通常在地面或灌木叢中覓食,以各種昆蟲和植物種子為食。它們的叫聲清亮悅耳,是森林中常聽到的聲音之一。
這些鳴禽不僅豐富了台灣的生物多樣性,也在生態系統中扮演著重要角色。它們通過捕食昆蟲來控制害蟲數量,通過傳播種子來促進植物的繁衍,有些還通過授粉來幫助植物繁殖。同時,這些鳴禽也是重要的環境指標,它們的數量和健康狀況能夠反映出生態系統的整體狀況。
3.18 台灣的夜行性鳥類
台灣最常見的夜行性鳥類之一—領角鴞(Otus lettia)。這種小型貓頭鷹以其特徵性的「耳朵」(實際上是羽毛)而得名。領角鴞主要生活在低海拔至中海拔的森林和農村地區,體長約20-25釐米。它們的羽毛呈褐色或灰色,有複雜的斑紋,非常有利於在樹林中隱蔽。領角鴞主要以各種昆蟲和小型脊椎動物為食,包括甲蟲、蛾類、小型哺乳動物和爬行動物。它們通常在樹洞或建築物的縫隙中築巢。領角鴞的叫聲是一種有節奏的「咕-咕」聲,在夜晚的森林中常能聽到。
黃嘴角鴞(Otus spilocephalus)是另一種在台灣常見的小型貓頭鷹。這種鳥比領角鴞稍小,體長約18-20釐米。如其名所示,黃嘴角鴞的喙呈黃色或淡綠色,這是它們的顯著特徵。它們的羽毛呈褐色或灰褐色,有複雜的斑點和條紋。黃嘴角鴞主要分佈在中高海拔的山區森林中,以昆蟲和小型脊椎動物為食。它們的叫聲是一種單調的「嗚」聲,在寂靜的山林中格外清晰。
褐鷹鴞(Ninox scutulata)是台灣體型最大的常見貓頭鷹。這種鳥體長約30-35釐米,羽毛以深褐色為主,胸部有淡褐色的縱紋。褐鷹鴞的眼睛呈黃色,非常明亮,是它們在夜間捕獵的重要工具。它們主要生活在低海拔至中海拔的森林中,食物包括中小型哺乳動物、鳥類、爬行動物和大型昆蟲。褐鷹鴞的叫聲是一種低沉的「呼-呼-呼」聲,在繁殖季節特別常聽到。
台灣夜鷹(Caprimulgus affinis)雖然不是貓頭鷹科的鳥類,但也是重要的夜行性鳥類。這種鳥屬於夜鷹科,體長約25-30釐米,羽毛呈褐色和灰色的複雜斑紋,非常有利於在地面或樹枝上隱蔽。台灣夜鷹有著特別寬大的嘴巴,非常適合在飛行中捕捉昆蟲。它們常在黃昏或夜晚時分飛行,發出特徵性的「咯-咯-咯」聲。台灣夜鷹主要分佈在低海拔的開闊地區,包括農田、草地和森林邊緣。
黑冠麻鷺(Gorsachius melanolophus)是一種夜行性鷺鳥,常見於台灣的低海拔森林和濕地周圍。這種鳥體長約50釐米,羽毛以褐色為主,頭頂有一塊黑色的羽冠。黑冠麻鷺在白天通常隱藏在茂密的植被中,到了夜晚則出來覓食。它們的食物主要包括魚類、兩棲類、甲殼類和水生昆蟲。黑冠麻鷺的叫聲是一種低沉的「呱-呱」聲,在夜晚的濕地周圍常能聽到。
夜鷺(Nycticorax nycticorax)是另一種在台灣常見的夜行性鷺鳥。這種鳥體型中等,成鳥的羽毛以灰色和白色為主,頭頂和背部呈黑色。夜鷺通常在黃昏時分開始活動,常常可以看到它們停在都市中的大樹或建築物上。它們主要以魚類和其他水生生物為食,因此在都市中的河流、湖泊或公園的水池附近常能看到它們的身影。夜鷺的叫聲是一種粗啞的「誇-誇」聲,在夜晚的城市中常能聽到。
3.19 台灣的稀有鳥類
台灣有許多稀有和特有的鳥類物種。以下是一些值得注意的例子:
1. 黑嘴端鳳頭燕鷗: 全球僅存約100隻,主要棲息在台灣的澎湖群島。
2. 台灣藍鵲: 台灣特有種,以其華麗的藍色羽毛和長尾巴聞名。
3. 黑面琵鷺: 全球瀕危物種,每年冬季會大量遷徙至台灣南部。
4. 綠島椋鳥: 僅生存於台灣東部的綠島上。
5. 台灣山鷓鴣: 台灣特有種,生活在中高海拔山區。
6. 黃山雀: 台灣特有亞種,棲息於高山森林。
7. 台灣畫眉: 台灣特有種,常見於中低海拔山區。
8. 林鵰: 在台灣屬於稀有猛禽,主要棲息於山區森林。
這些鳥類不僅對台灣的生態系統很重要,也是台灣生物多樣性的重要指標。保護這些稀有物種及其棲息地對維護台灣的自然遺產至關重要。
您是否想瞭解更多關於這些鳥類或台灣鳥類保育的資訊
3.20 台灣的外來入侵種鳥類
台灣確實面臨一些外來入侵鳥類的問題。這些物種可能對本地生態系統造成威脅。以下是一些在台灣的主要外來入侵鳥類:
1. 白尾八哥:原產於南亞,現在在台灣許多地區常見。
2. 家八哥:也稱為爪哇八哥,原產於東南亞。
3. 綠繡眼:原產於亞洲其他地區,現已在台灣廣泛分佈。
4. 埃及聖䴉:原產於非洲,在台灣的族群正在擴大。
5. 紅嘴藍鵲:原產於中國大陸,在台灣部分地區有野生族群。
6. 鵲鴝:原產於中國大陸,在台灣中部有野生族群。
7. 橙嘴烏鴉:原產於東南亞,在台灣南部有發現。
8. 斑胸草雀:原產於非洲和亞洲其他地區,在台灣部分地區有野生族群。
這些外來種可能會與本地鳥類競爭資源,有些甚至可能捕食本地物種。此外,它們可能攜帶疾病,對農業造成損害,或改變當地生態平衡。
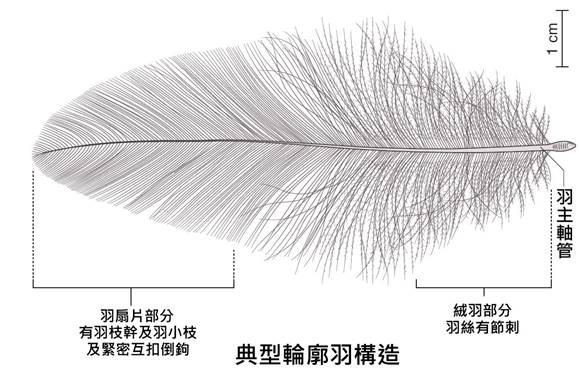
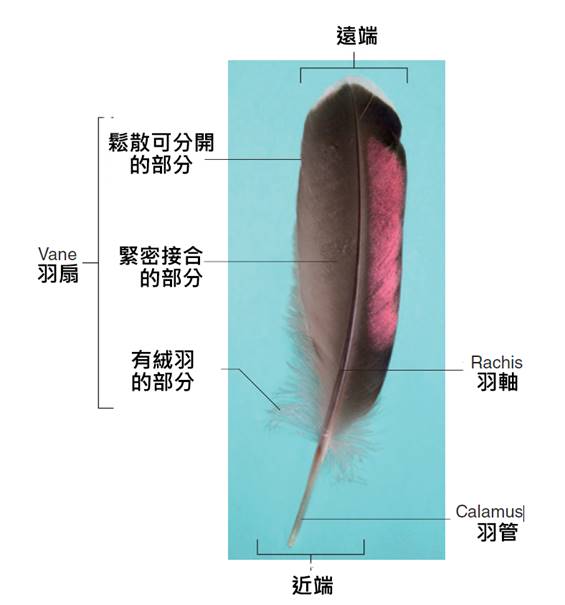
第四章 羽毛與羽衣

4.1羽毛的結構與發育
羽毛是鳥類最為獨特和顯著的特徵之一,其結構複雜而精巧,發育過程更是一個精密的生物學奇蹟。羽毛不僅賦予了鳥類飛行的能力,還在保溫、防水、求偶展示等多個方面發揮著關鍵作用。深入瞭解羽毛的結構與發育,有助於我們更全面地認識鳥類的生理特性和生態適應。
羽毛的基本結構包括幾個主要部分:羽軸、羽支和羽小枝。羽軸是羽毛的中心軸,由兩部分組成:位於皮膚內的根部稱為羽根,暴露在外的部分稱為羽幹。羽幹是羽毛的主要支撐結構,通常是中空的,這種設計既保證了強度,又減輕了重量。從羽幹的兩側延伸出的扁平結構稱為羽支,羽支上再分支出更細小的羽小枝。在大多數飛羽中,羽小枝上還有微小的鉤狀結構,這些鉤狀結構能夠相互咬合,形成一個平滑、緊密的羽片表面,這對於飛行至關重要。
羽毛的發育是一個複雜的過程,始於胚胎期的皮膚。在鳥胚的皮膚表面,會形成規律排列的羽毛原基。這些原基是由表皮和真皮共同參與形成的小突起。隨著發育的進行,羽毛原基會逐漸下陷形成羽毛濾泡。在濾泡的底部,一群快速分裂的細胞形成了羽毛芽。羽毛芽中的細胞會進行複雜的分化和排列,最終形成羽毛的各個組成部分。
羽毛的生長是一個連續的過程。新的羽毛在濾泡中形成並生長,逐漸推出舊的羽毛。這個過程通常在換羽期間發生,但某些鳥類物種可能全年都在不斷更新羽毛。羽毛生長的速度可以非常快,某些大型鳥類的飛羽每天可以生長超過1釐米。
羽毛的發育過程中,不同類型的蛋白質扮演著重要角色。角蛋白是羽毛的主要成分,它提供了強度和彈性。另外,一些特殊的蛋白質負責產生羽毛的顏色。例如,黑色素細胞產生的黑色素可以給羽毛賦予黑色或棕色。而某些蛋白質的特殊排列方式可以產生結構性顏色,如藍色或綠色。
羽毛的微觀結構決定了其特定的功能。例如,飛羽的羽小枝緊密咬合,形成平滑的表面,這對於產生升力至關重要。而絨羽的結構則更為鬆散,羽小枝之間沒有鉤狀結構,這種設計能夠有效地困住空氣,提供excellent的保溫效果。
有趣的是,羽毛的發育過程在某種程度上重現了羽毛的演化歷史。最原始的羽毛可能只是簡單的絲狀結構,類似於現代鳥類的絨羽。隨後,這些結構逐漸演化出分支,最終形成了具有複雜羽支和羽小枝結構的現代羽毛。這種發育過程與演化歷史的對應關係,為我們理解羽毛如何從簡單的保溫結構演變為複雜的飛行器官提供了重要線索。
羽毛的生長受到多種因素的影響,包括營養狀況、激素水準和環境條件等。營養不良可能導致羽毛生長不良或顏色淡化。某些激素,如甲狀腺素和性激素,也會影響羽毛的生長和著色。環境因素如光照週期,則可能影響換羽的時間和進程。
羽毛的發育還涉及複雜的基因調控網絡。近年來的研究發現,多個基因家族參與了羽毛的形成和發育。例如,(BMP)和(Shh)信號通路在羽毛原基的形成和羽支的分支中起重要作用。而(Keratin)基因家族則負責產生構成羽毛主體的角蛋白。瞭解這些基因的功能和調控機制,不僅有助於我們理解羽毛發育的分子機制,還為研究羽毛的演化提供了新的視角。
羽毛的生長是一個高度能量消耗的過程。在換羽期間,鳥類的能量需求會顯著增加。為了滿足這一需求,許多鳥類會改變其行為模式,例如減少活動或增加進食。一些遷徙鳥類甚至會根據換羽的需要調整其遷徙時間表。這種現象突顯了羽毛對鳥類生存的重要性,同時也展示了鳥類如何平衡不同生理需求。
值得注意的是,雖然所有鳥類的羽毛都遵循相似的基本發育模式,但不同物種的羽毛可能表現出顯著的差異。這些差異可能體現在羽毛的大小、形狀、顏色或特殊結構上。例如,某些天堂鳥物種的裝飾性羽毛可以長到極其誇張的程度,而企鵝的羽毛則演化出了特殊的防水結構。這種多樣性反映了羽毛如何通過演化適應不同的生態需求和性選擇壓力。
羽毛的發育不僅限於胚胎期和幼年期,成年鳥類也會定期更新羽毛。這個過程稱為換羽,是羽毛發育的一個重要方面。換羽模式因物種而異,可能是全身同時進行,也可能是逐步更換。某些鳥類一年只換羽一次,而另一些則可能一年換羽多次。換羽過程的精確控制對於鳥類至關重要,因為它不僅影響外觀,還直接關係到飛行能力和保溫效果。
羽毛的發育還與鳥類的年齡和性成熟有關。許多鳥類物種的幼鳥羽衣與成鳥有明顯區別,這種差異可能有助於幼鳥在成長期間保持隱蔽。隨著鳥類逐漸成熟,羽衣會經歷一系列變化,最終達到成年羽衣。在某些物種中,性成熟也會引發羽衣的顯著變化,特別是在存在性二態的物種中。
羽毛的結構和發育過程也為生物材料科學提供了靈感。研究人員正在研究如何模仿羽毛的微觀結構來開發新型輕質、高強度材料。同時,瞭解羽毛的生長機制也可能為治療皮膚疾病或促進毛髮生長提供新的思路。
深入研究羽毛的結構與發育不僅有助於我們更好地理解鳥類生物學,還為進化生物學、生態學和生物材料科學等領域提供了豐富的研究素材。隨著研究技術的不斷進步,我們對羽毛這一神奇結構的認識也在不斷深化,揭示出更多關於鳥類適應性和生物多樣性的奧秘。
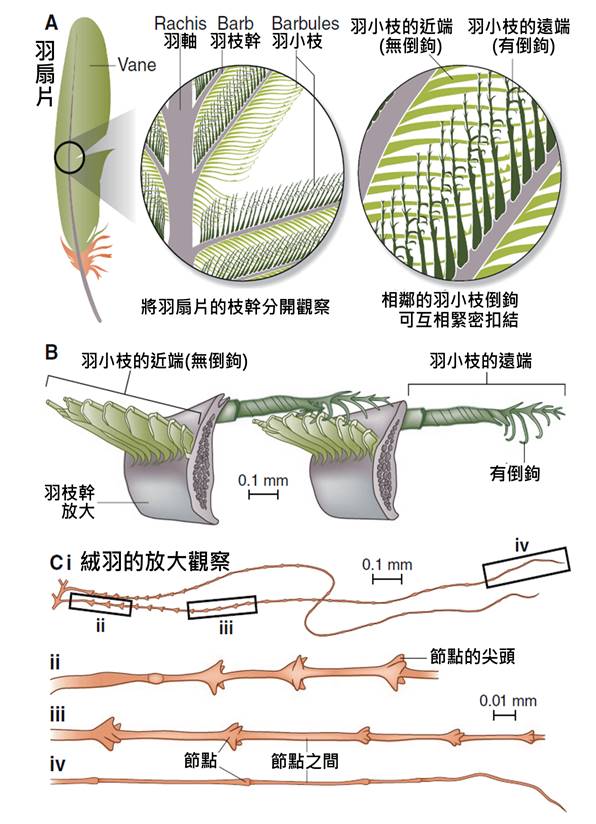

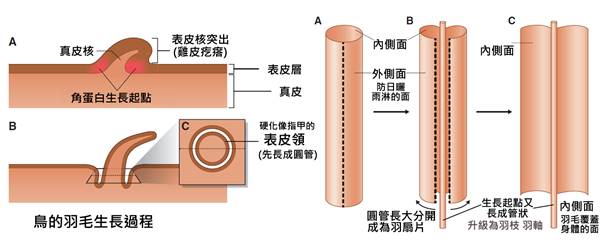
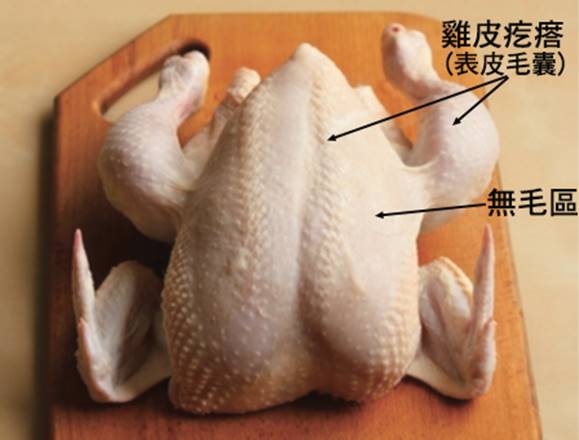
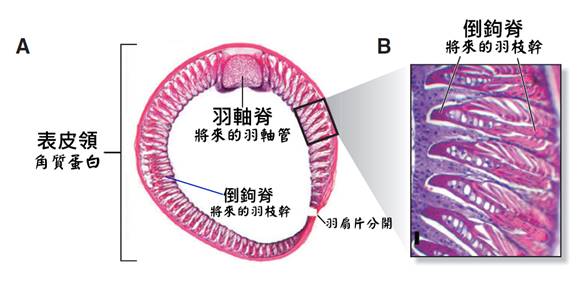
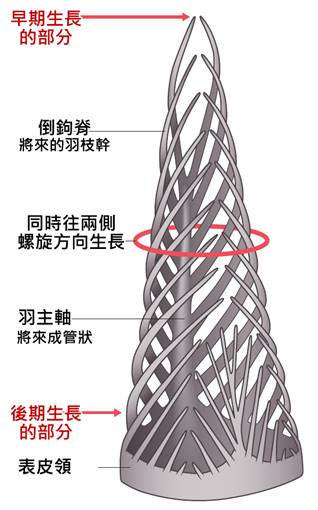
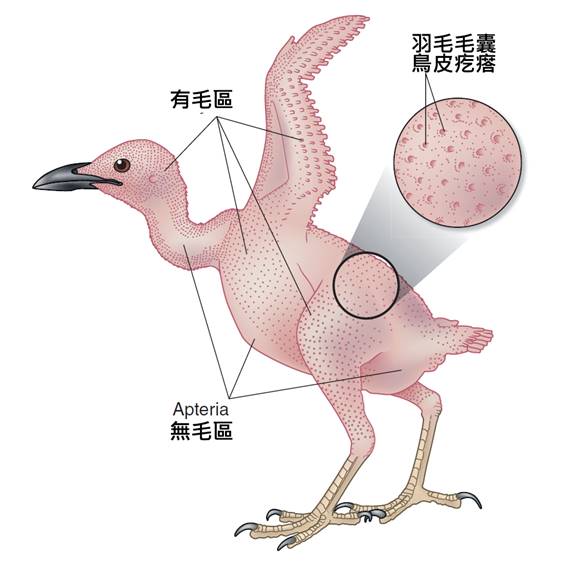
4.2羽毛的類型與功能
鳥類的羽毛是一種獨特而複雜的結構,在鳥類的生存和適應中扮演著至關重要的角色。羽毛不僅僅是鳥類外表的裝飾,更是其生理功能和行為表現的關鍵要素。鳥類的羽毛種類繁多,每種羽毛都有其特定的功能和用途。
首先,我們來談談最常見的羽毛類型—輪廓羽(Contourfeathers)。這種羽毛覆蓋了鳥類身體的大部分表面,形成了鳥類特有的流線型外形。輪廓羽具有堅固的羽軸和緊密的羽片結構,能夠為鳥類提供保護,抵禦外界環境的影響。同時,輪廓羽還具有良好的防水性,這對於水鳥尤為重要。在飛行過程中,輪廓羽能夠幫助鳥類保持身體的平衡和穩定,減少空氣阻力。
翼羽(Flightfeathers)是另一種極為重要的羽毛類型,主要分佈在鳥類的翅膀和尾部。翼羽包括初級飛羽、次級飛羽和尾羽。初級飛羽位於翅膀的外側,是推動鳥類飛行的主要力量來源。次級飛羽位於翅膀的內側,主要用於提供升力。尾羽則幫助鳥類在飛行中進行轉向和平衡。這些特化的羽毛結構使得鳥類能夠在空中自如地飛翔,展現出令人驚嘆的飛行技巧。
絨羽(Downfeathers)是一種柔軟蓬鬆的羽毛,主要分佈在鳥類的皮膚表面,被輪廓羽覆蓋。絨羽的主要功能是保溫,它們能夠在羽毛之間形成大量的空氣隔層,有效地阻隔外界的冷空氣,維持鳥類的體溫。在幼鳥身上,絨羽尤為重要,因為幼鳥還沒有發育出完整的輪廓羽。
半絨羽(Semiplumefeathers)是介於輪廓羽和絨羽之間的一種羽毛類型。它們具有羽軸,但羽片結構較為鬆散。半絨羽的主要功能是填充鳥類身體的輪廓,同時也具有一定的保溫作用。
纖羽(Filoplumefeathers)是一種極細小的羽毛,通常分佈在其他羽毛的周圍。纖羽的末端有感覺神經,能夠感知其他羽毛的位置和運動狀態,幫助鳥類調整羽毛的位置,以達到最佳的飛行狀態。
刷毛羽(Bristlefeathers)是一種特殊的羽毛類型,通常分佈在鳥類的頭部,特別是嘴部和眼睛周圍。這種羽毛的羽軸堅硬,但幾乎沒有羽片。刷毛羽的主要功能是保護鳥類的眼睛和鼻孔,防止灰塵和其他小顆粒進入。對於一些捕食昆蟲的鳥類來說,刷毛羽還可以幫助它們感知和捕捉獵物。
粉羽(Powderdownfeathers)是一種特殊的羽毛,只存在於某些鳥類品種中,如鸚鵡和蒼鷺。這種羽毛會持續生長並在末端分解成細小的粉末。這些粉末具有防水和清潔的作用,鳥類會將其散佈到其他羽毛上,幫助維護羽毛的整潔和功能。
除了上述主要的羽毛類型,還有一些特殊的羽毛結構值得一提。例如,一些鳥類的尾羽演化出了特殊的形態,如孔雀的華麗尾屏,主要用於求偶展示。另外,有些鳥類的翼羽邊緣具有鋸齒狀結構,可以減少飛行時產生的噪音,這對於貓頭鷹等夜行性捕食者特別重要。
羽毛的顏色也是一個值得關注的方面。鳥類羽毛的色彩可以由色素決定,也可能是由羽毛的微觀結構造成的結構色。色素如黑色素和類胡蘿蔔素可以直接賦予羽毛顏色,而結構色則是由羽毛表面的微觀結構反射特定波長的光線而產生的。這些豐富多彩的羽毛不僅用於求偶和種內識別,還可能具有保護色的功能,幫助鳥類在環境中隱藏自己。
羽毛的生長和更換也是鳥類生理週期中的重要部分。大多數鳥類每年都會經歷換羽過程,更換舊的或損壞的羽毛。這個過程通常是漸進的,以確保鳥類在換羽期間仍能保持飛行能力。然而,有些水鳥物種會在短時間內更換所有飛羽,導致暫時失去飛行能力。
羽毛還具有重要的社會和行為功能。許多鳥類會通過豎起或放下某些部位的羽毛來傳遞資訊,例如表達威脅、求偶或臣服。一些鳥類還會利用羽毛產生聲音,如啄木鳥用尾羽支撐身體,或者燕雀科鳥類用翅膀發出嗡嗡聲以吸引異性。
在進化的長河中,羽毛的功能不斷擴展和多樣化。最初可能僅用於保溫,後來演化出了飛行功能,並進一步發展出各種特殊用途。這種多功能性使得羽毛成為鳥類適應各種生態環境的關鍵因素。
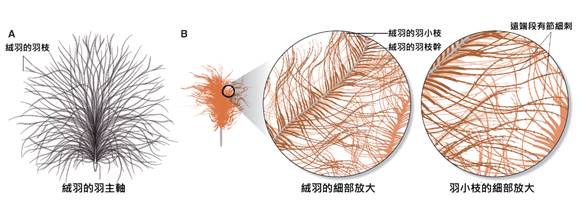
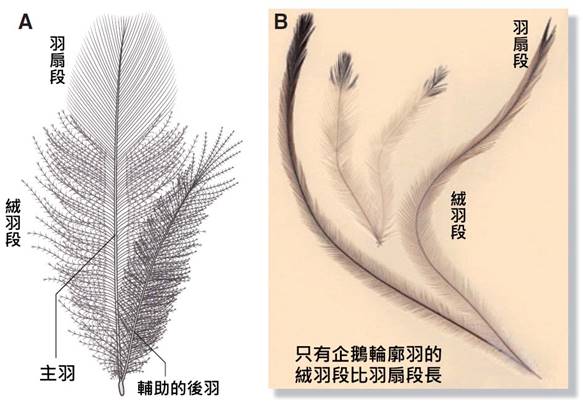
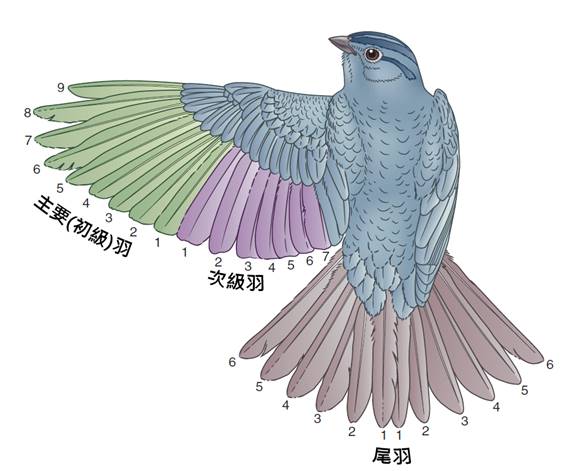
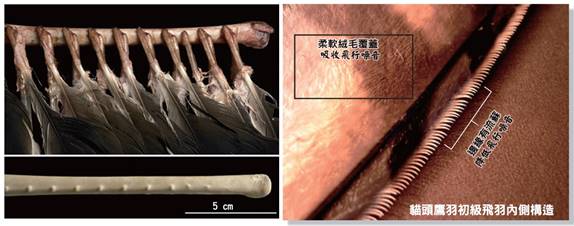

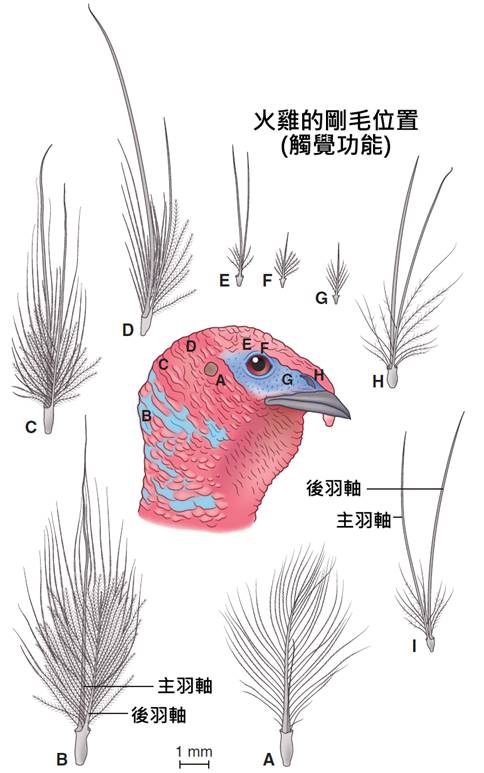


4.3換羽與羽衣變化
換羽是鳥類生命週期中一個至關重要的生理過程,這個過程涉及舊羽毛的脫落和新羽毛的生長。對於鳥類而言,換羽不僅僅是更新外表,更是維持生存能力的必要條件。羽毛會隨著時間的推移而磨損、褪色或受損,因此定期更換羽毛對於保持飛行能力、保溫效果以及整體健康至關重要。
鳥類的換羽模式因物種而異,但通常可以分為兩種主要類型:完全換羽和部分換羽。完全換羽指的是鳥類在一個特定的時期更換所有羽毛,這通常發生在繁殖季節結束後。部分換羽則是只更換部分羽毛,這種模式在許多鳥類中更為常見,特別是在年輕鳥類和成年鳥類之間的過渡期。
換羽過程通常是漸進的,以確保鳥類在此期間仍能保持基本的飛行能力和體溫調節。然而,有些水鳥種類,如鴨子和天鵝,會同時脫落所有飛羽,導致暫時失去飛行能力。這種被稱為「同步換羽」的現象通常發生在安全的水域環境中,鳥類可以在水中躲避天敵,直到新的飛羽長出。
換羽的時間和頻率受多種因素影響,包括鳥類的年齡、性別、繁殖狀態以及環境條件。大多數成年鳥類每年至少換羽一次,通常在繁殖季節結束後。一些物種甚至每年換羽兩次,分別在春季和秋季。年輕鳥類可能在第一年內經歷多次部分換羽,直到獲得成年羽衣。
換羽過程對鳥類來說是一項巨大的生理挑戰,需要消耗大量能量。新羽毛的生長需要大量蛋白質和其他營養物質,因此鳥類在換羽期間通常會增加食物攝入量。一些鳥類甚至會改變其行為模式,減少活動以節省能量。換羽過程中,鳥類可能會顯得較為虛弱和易受傷害,因此許多鳥類會選擇在食物豐富、天敵威脅較小的時期和地點進行換羽。
羽衣變化是與換羽密切相關的另一個重要現象。羽衣指的是鳥類在不同生命階段或季節所呈現的整體外觀。羽衣變化可以反映鳥類的年齡、性別、繁殖狀態以及季節性適應。
最常見的羽衣變化類型包括年齡相關的變化、季節性變化和性別二態性。年齡相關的羽衣變化通常發生在幼鳥向成鳥過渡的過程中。幼鳥的羽衣往往較為樸素,有助於在巢中或離巢初期的保護色。隨著年齡增長,鳥類會逐漸獲得成年羽衣,這通常需要經過多次換羽。
季節性羽衣變化在許多鳥類中很常見,尤其是在溫帶地區。這種變化通常與繁殖季節相關。在繁殖季節,許多鳥類會換上更鮮豔、更醒目的羽衣,這被稱為繁殖羽。繁殖羽通常更為華麗,有助於吸引配偶和宣示領地。相比之下,非繁殖季節的羽衣(也稱為冬羽或基本羽)通常較為樸素,有助於鳥類在環境中隱藏自己,避免天敵的注意。
性別二態性指的是雄鳥和雌鳥在外觀上的差異,這在許多鳥類物種中很常見。通常,雄鳥的羽衣更為鮮豔醒目,特別是在繁殖季節,而雌鳥的羽衣則較為樸素。這種差異可能是由性選擇驅動的進化結果,鮮豔的雄鳥更容易吸引雌鳥,而樸素的雌鳥則更容易在孵卵和育雛時隱藏自己。然而,並非所有鳥類都表現出明顯的性別二態性,有些物種雌雄外觀相似。
值得注意的是,一些鳥類物種可以通過部分換羽快速改變其外觀,而無需完全更換羽毛。這種現象被稱為「磨損換羽」。例如,一些雀形目鳥類在冬季時,其羽毛的尖端較暗或有不同的顏色。隨著春季的到來,這些羽毛尖端會自然磨損脫落,露出下面更鮮豔的羽毛,使鳥類呈現出繁殖季節的外觀,而無需進行完整的換羽。
羽衣變化還可能受到環境因素的影響。例如,一些極地鳥類會在冬季換上白色羽衣以適應雪地環境,而在夏季則換回較深色的羽衣。這種適應性變化有助於鳥類在不同季節的環境中更好地偽裝自己。
此外,羽衣的狀態可以反映鳥類的健康狀況和整體適應度。健康的鳥類通常有光滑、緊密的羽毛,顏色鮮豔。相反,營養不良、患病或受到寄生蟲侵擾的鳥類可能會表現出羽毛質地粗糙、顏色暗淡等症狀。因此,觀察鳥類的羽衣狀態對於研究人員和鳥類愛好者來說是一個重要的健康指標。
在進化的角度來看,換羽和羽衣變化是鳥類適應不同環境和生存挑戰的重要機制。通過調整羽衣的顏色、密度和結構,鳥類能夠在不同的季節和生態環境中優化其生存策略。這種適應性不僅體現在個體層面,也反映了物種層面的進化適應。
換羽和羽衣變化的研究對於鳥類學和生態學具有重要意義。通過研究這些現象,科學家們可以更好地瞭解鳥類的生活史策略、遷徙模式、繁殖行為以及對環境變化的響應。這些知識不僅有助於我們更好地理解和保護鳥類,還為研究生物適應性和進化提供了寶貴的見解。

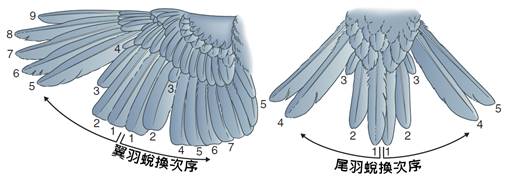
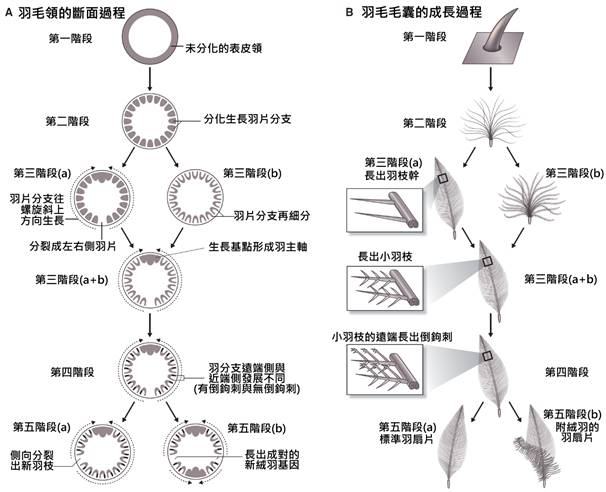
4.4羽毛的顏色與紋理
鳥類羽毛的顏色和紋理是自然界中最引人注目的現象之一,不僅展現了生物多樣性的魅力,也反映了鳥類在進化過程中的適應策略。羽毛的顏色和紋理不僅僅是美學上的裝飾,更在鳥類的生存、繁衍和社交中扮演著至關重要的角色。
羽毛的顏色主要源於兩種機制:色素沉積和結構色。色素是最常見的羽毛著色方式,主要包括黑色素、類胡蘿蔔素和卟啉。黑色素主要產生黑色、灰色和褐色等深色調,是鳥類羽毛中最普遍的色素。黑色素不僅賦予羽毛顏色,還能增強羽毛的強度和耐磨性。類胡蘿蔔素則負責產生黃色、橙色和紅色等鮮豔色彩。有趣的是,鳥類無法自身合成類胡蘿蔔素,必須通過食物攝入,因此類胡蘿蔔素的含量某種程度上反映了鳥類的營養狀況和覓食能力。卟啉色素較為罕見,主要產生紅色和綠色,在某些鳥類如鵜鶘的羽毛中可以觀察到。
結構色是另一種令人著迷的羽毛著色機制,它不依賴於色素,而是通過羽毛的微觀結構對光線進行散射、折射和干涉而產生的。結構色通常呈現出金屬光澤或彩虹般的光變效果,如孔雀羽毛的鮮豔藍綠色和蜂鳥羽毛的閃耀色彩。結構色的形成主要依賴於羽毛中的角蛋白結構,這些結構形成了納米級的週期性排列,能夠選擇性地反射特定波長的光線。結構色的一個顯著特點是其顏色會隨觀察角度的變化而改變,這種現象被稱為「閃色」(Iridescence)。
除了單一的色素或結構色,許多鳥類的羽毛顏色是由多種機制共同作用產生的。例如,綠色羽毛通常是黃色類胡蘿蔔素與藍色結構色相結合的結果。這種複雜的顏色形成機制使得鳥類能夠展現出豐富多樣的色彩,從而適應不同的生態環境和社交需求。
羽毛的紋理是另一個重要的視覺特徵,它與顏色相輔相成,共同構成了鳥類獨特的外觀。紋理可以表現為斑點、條紋、環帶或複雜的幾何圖案。這些紋理不僅增加了視覺上的複雜性和美感,還具有重要的生物學功能。例如,許多地棲鳥類的羽毛上的斑點和條紋可以提供excellent的偽裝效果,幫助它們融入背景環境,避免被天敵發現。相反,一些鳥類的鮮明紋理則用於種內識別、求偶展示或警告信號。
羽毛顏色和紋理的形成受到多種因素的影響,包括遺傳、環境和生理狀況。遺傳因素決定了鳥類產生特定色素或形成特定微觀結構的能力。環境因素,如食物來源、氣候條件和棲息地特徵,可以影響色素的沉積和羽毛結構的發育。生理狀況,包括年齡、性別、健康狀態和激素水準,也會對羽毛的顏色和紋理產生顯著影響。
在許多鳥類物種中,雄性和雌性的羽毛顏色和紋理存在顯著差異,這種現象被稱為性別二態性。通常,雄性會有更鮮豔、更複雜的羽毛圖案,這主要是由性選擇驅動的。鮮豔的羽毛可以作為雄性健康、活力和基因質量的指標,有助於吸引雌性。相比之下,雌性的羽毛往往較為樸素,這有助於在孵卵和育雛期間提供保護色。然而,這種模式並非普遍適用,有些物種中雌性反而更為鮮豔,或者雌雄羽色相似。
羽毛的顏色和紋理還可能隨季節變化而改變,這種現象被稱為季節性二態性。許多鳥類在繁殖季節會換上更鮮豔的羽衣,而在非繁殖季節則換回較為樸素的羽衣。這種變化不僅涉及顏色,還可能包括特殊的裝飾性羽毛的生長或脫落。季節性的羽衣變化使鳥類能夠在不同的季節中優化其外觀,以適應繁殖、遷徙或越冬等不同的生活階段需求。
值得注意的是,羽毛的顏色和紋理不僅影響鳥類的視覺外觀,還可能對其他功能產生影響。例如,深色羽毛通常能夠更有效地吸收陽光,有助於體溫調節。某些紋理可能影響羽毛的空氣動力學特性,從而影響飛行效率。此外,一些鳥類的特殊羽毛結構還可以產生聲音,用於通訊或求偶。
在生態學和進化生物學的背景下,羽毛的顏色和紋理提供了豐富的研究素材。通過研究不同物種的羽毛特徵,科學家們可以探索物種間的親緣關係、適應性輻射過程以及環境壓力對物種演化的影響。例如,棲息在相似環境中的不相關物種可能演化出相似的羽毛顏色和紋理,這種現象被稱為趨同進化。
羽毛顏色和紋理的研究還涉及了複雜的物理和化學過程。例如,結構色的形成機制涉及光學和材料科學的原理,而色素的合成和沉積則涉及生物化學和分子生物學的知識。這種跨學科的研究方法不僅加深了我們對鳥類羽毛的理解,還為生物材料的開發和仿生技術的應用提供了靈感。
在保護生物學的角度來看,羽毛的顏色和紋理可以作為評估種群健康和環境質量的指標。環境污染、氣候變化和棲息地破壞等因素可能會影響鳥類羽毛的顏色表現,從而反映生態系統的整體健康狀況。因此,長期監測鳥類羽毛的顏色和紋理變化可以為生態保護工作提供重要的參考資訊。
羽毛的顏色和紋理不僅是自然之美的體現,更是鳥類適應性進化的結果,反映了物種與環境之間複雜而精妙的相互作用。通過深入研究這一領域,我們不僅可以更好地理解和欣賞自然界的多樣性,還能為生物技術創新和生態保護提供新的思路和方法。

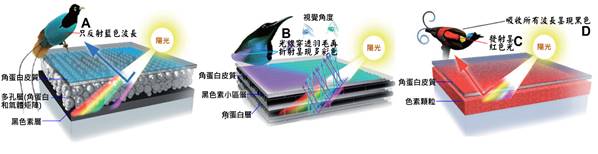

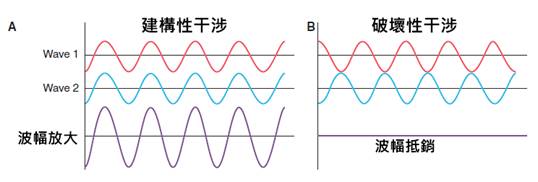
4.5羽毛的視覺功能
鳥類的羽毛不僅是飛行和保溫的重要工具,還在視覺交流和信號傳遞方面扮演著至關重要的角色。羽毛的視覺功能涉及多個方面,包括種內識別、求偶展示、警戒信號、偽裝保護等,這些功能共同構成了鳥類複雜而豐富的視覺交流系統。
在種內識別方面,羽毛的顏色和紋理提供了重要的視覺線索。許多鳥類物種通過特定的羽毛特徵來識別同類,這不僅包括整體的羽色模式,還可能涉及特定部位的獨特標記。例如,某些海鳥物種的頭部或翅膀上可能有獨特的色斑,這些標記可以在遠距離或大型群落中快速識別個體。這種識別機制在繁殖季節尤為重要,有助於鳥類找到合適的配偶並維護領地。
求偶展示是羽毛視覺功能的另一個重要方面。在許多鳥類物種中,雄性通常擁有更鮮豔、更華麗的羽毛,這些羽毛在求偶期間扮演著關鍵角色。例如,孔雀的雄鳥擁有令人驚嘆的尾屏,在求偶時展開,形成壯觀的視覺效果。這種誇張的羽毛裝飾不僅吸引雌性的注意,還可能傳達雄性的健康狀況和基因質量。有趣的是,一些研究表明,羽毛的對稱性和色彩飽和度可能與個體的整體適應度相關,因此雌性通過評估這些視覺特徵來選擇最佳的配偶。
警戒信號是羽毛視覺功能的另一個重要方面。一些鳥類物種擁有特殊的羽毛結構或顏色模式,在感受到威脅時可以快速展示。例如,某些猛禽種類在受到驚擾時會豎起頭頂的羽毛,形成醒目的冠羽,這種視覺信號可以威懾潛在的敵人或競爭對手。同樣,一些地棲鳥類在飛行時會露出隱藏的鮮艷羽毛,這種突然的色彩變化可以驚嚇捕食者,為逃生爭取寶貴的時間。
偽裝保護是羽毛視覺功能中極為重要的一環。通過特定的羽毛顏色和紋理,許多鳥類能夠完美地融入其棲息環境。例如,沙漠地區的鳥類通常擁有沙褐色的羽毛,而森林中的鳥類可能具有斑駁的綠色和棕色羽毛,這些顏色和紋理使它們能夠有效地隱藏自己,避免被天敵發現。一些鳥類甚至能夠根據季節變化調整羽毛顏色,如雷鳥在冬季會換上白色羽毛以適應雪地環境。這種適應性偽裝不僅保護了鳥類免受捕食,還有助於它們在捕獵時接近獵物。
羽毛的視覺功能還涉及一些更為微妙的方面。例如,紫外線反射在鳥類的視覺交流中扮演著重要角色。與人類不同,許多鳥類物種能夠感知紫外光。一些看似單調的羽毛在紫外光下可能呈現出複雜的圖案,這些圖案可能用於種內識別或者性別辨認。這種「隱藏」的視覺信號為鳥類提供了一種人類無法直接觀察到的交流方式。
羽毛的結構色也是視覺功能的重要組成部分。結構色不依賴於色素,而是由羽毛的微觀結構產生,這種顏色往往呈現出金屬光澤或者閃耀效果。結構色的一個顯著特點是其顏色會隨觀察角度的變化而改變,這種動態的視覺效果在某些物種的求偶展示中起到了關鍵作用。例如,蜂鳥的喉部羽毛就展現出令人驚嘆的閃耀效果,這種效果不僅吸引了配偶的注意,還可能傳達個體質量的資訊。
羽毛的視覺功能還與鳥類的行為緊密相連。許多鳥類物種會通過特定的姿態或動作來強化羽毛的視覺效果。例如,一些雀形目鳥類在領地爭奪或求偶時會豎起頭冠羽毛,這不僅增加了身體的視覺大小,還展示了特定的顏色模式。同樣,一些涉禽在警戒時會迅速展開尾羽,露出隱藏的鮮艷顏色,這種突然的視覺變化可以有效地警告群體中的其他成員。
在群體生活的鳥類中,羽毛的視覺功能還扮演著維持社會結構的重要角色。例如,在某些鸚鵡物種中,個體的羽色可能反映其在群體中的地位。dominant個體可能擁有更鮮豔或更完整的羽色,這種視覺信號有助於減少群體內的不必要衝突,維持穩定的社會階層。
季節性的羽毛變化也是視覺功能的一個重要方面。許多鳥類物種會在繁殖季節和非繁殖季節之間轉換羽色。這種變化不僅適應了不同季節的環境需求(如保護色的調整),還可能反映個體的生理狀態和繁殖準備程度。例如,一些雀形目鳥類在繁殖季節會獲得更鮮艷的羽色,這不僅是求偶信號,還可能表明個體已經生理成熟,準備好進行繁殖。
羽毛的視覺功能還在種間互動中發揮作用。某些鳥類物種可能進化出模仿其他物種羽色的能力,這種擬態可能用於欺騙捕食者或者獲得某些生態優勢。例如,某些杜鵑科的鳥類會模仿猛禽的羽色,以嚇阻潛在的捕食者。
在生態系統中,羽毛的視覺功能還可能影響鳥類的分佈和行為模式。例如,在密林環境中,鮮豔的羽毛可能更容易被捕食者發現,因此這樣的環境可能促進了較為樸素羽色的進化。相反,在開闊地帶,鮮豔的羽毛可能在遠距離交流中更為有效,因此可能favors更鮮艷羽色的演化。
羽毛的視覺功能還為科學研究提供了豐富的素材。通過研究不同物種的羽毛特徵,科學家們可以探索物種間的演化關係、生態適應過程以及行為生態學的諸多問題。例如,通過比較近緣物種的羽毛特徵,研究人員可以推斷環境因素如何塑造了物種的視覺信號系統。
羽毛的視覺功能不僅限於鳥類之間的交流,還影響了鳥類與其他生物的互動。例如,某些授粉鳥類的羽毛顏色可能與它們訪問的花朵顏色相協調,這種共同進化的結果不僅有利於鳥類獲取花蜜,還促進了植物的授粉過程。
4.6羽毛的護理
羽毛護理是鳥類生活中不可或缺的重要部分,直接關係到其生存、繁衍和整體健康。鳥類投入大量時間和精力來維護羽毛的狀態,這不僅僅是為了保持外表美觀,更是為了確保羽毛能夠充分發揮其多重功能,包括飛行、保溫、防水和視覺信號等。
鳥類的羽毛護理行為主要包括梳理、整理、清潔和上油等多個方面。這些行為不僅是鳥類日常生活的重要組成部分,也反映了鳥類在長期進化過程中形成的適應性策略。
梳理是最常見的羽毛護理行為之一。鳥類會使用喙來梳理全身的羽毛,這個過程不僅能夠清理羽毛上的灰塵和雜物,還能夠重新排列羽片,使羽毛恢復正確的形狀和結構。在梳理過程中,鳥類會特別注意飛羽和尾羽,因為這些羽毛對飛行至關重要。通過仔細地梳理每一根羽毛,鳥類能夠確保羽片之間緊密連接,形成光滑的表面,從而優化飛行性能。
清潔是另一個重要的羽毛護理環節。鳥類採用多種方法來清潔羽毛,其中最常見的是水浴。許多鳥類會定期在淺水中沐浴,通過浸泡和拍打羽毛來去除污垢和寄生蟲。水浴不僅能夠清潔羽毛,還能夠幫助鳥類調節體溫。一些生活在乾旱地區的鳥類則會進行塵浴,通過在乾燥的沙土或灰塵中翻滾來清潔羽毛。塵浴能夠吸收羽毛上的多餘油脂,並有助於去除寄生蟲。
上油是羽毛護理中一個特別有趣的行為。大多數鳥類的尾部上方都有一個稱為尾脂腺(Uropygialgland)的特殊腺體,這個腺體分泌油脂。鳥類會用喙擠壓尾脂腺,然後將分泌的油脂塗抹到全身羽毛上。這種油脂具有多重功能:首先,它能夠使羽毛保持柔軟和彈性,防止羽毛變脆和斷裂;其次,油脂能夠形成一層防水膜,這對水鳥尤為重要,能夠防止羽毛被水浸透;再者,油脂還具有一定的抗菌和防黴效果,有助於維護羽毛的健康;最後,一些研究表明,尾脂腺分泌物中可能含有能夠吸引異性的資訊素,在鳥類的社交和繁殖中可能起到重要作用。
羽毛的護理還包括定期的換羽過程。換羽是鳥類更新和替換舊羽毛的自然過程,通常每年發生一次或兩次。在換羽期間,鳥類會逐漸脫落舊羽毛,同時生長新羽毛。這個過程需要消耗大量能量,因此鳥類在換羽期間通常會減少活動,增加食物攝入。換羽不僅能夠更新受損或磨損的羽毛,還能夠調整羽毛的顏色和結構,以適應季節性的需求,如繁殖期的華麗羽衣或冬季的保暖羽毛。
在羽毛護理過程中,鳥類還會特別注意某些特定部位的羽毛。例如,飛行羽毛(包括初級飛羽和次級飛羽)對於飛行能力至關重要,因此鳥類會格外小心地護理這些羽毛。同樣,頭部和頸部的羽毛也受到特別關注,因為這些部位的羽毛對於保護重要器官和維持體溫起著關鍵作用。
羽毛護理行為還與鳥類的社交生活密切相關。許多鳥類物種會進行互相梳理(Allopreening)的行為,即個體之間互相幫助清理和整理羽毛。這種行為不僅有助於清理難以自己觸及的部位,如頭部和頸部,還能夠加強個體之間的社會聯繫。在一些物種中,互相梳理還可能是求偶或鞏固配對關係的重要行為。
環境因素對羽毛護理也有重要影響。例如,生活在污染嚴重地區的鳥類可能需要更頻繁地清潔羽毛,以去除沉積在羽毛上的污染物。同樣,棲息在寄生蟲豐富環境中的鳥類可能會更頻繁地進行塵浴或陽光浴,以控制外部寄生蟲的數量。氣候條件也會影響羽毛護理行為,例如,在乾燥氣候中生活的鳥類可能需要更頻繁地進行上油行為,以防止羽毛變得乾燥和脆弱。
羽毛護理還涉及一些特殊的行為適應。例如,一些水鳥物種能夠將頭部完全轉向後方,這種靈活性使它們能夠更有效地清理和上油背部的羽毛。另一個有趣的例子是,某些鸚鵡物種會使用樹皮或葉子等天然物質來「刷洗」羽毛,這種行為可能有助於去除寄生蟲或調節羽毛上的油脂含量。
在某些情況下,羽毛護理行為可能會受到限制或改變。例如,受傷或生病的鳥類可能無法正常進行羽毛護理,這可能導致羽毛狀況迅速惡化。同樣,處於高度壓力下的鳥類可能會減少羽毛護理行為,將精力集中在生存和警戒上。在人工飼養環境中,如動物園或救護中心,工作人員需要特別關注鳥類的羽毛護理情況,有時甚至需要人工協助進行羽毛護理。
羽毛護理的重要性還體現在鳥類的遷徙行為中。長途遷徙的鳥類在啟程前會花費大量時間進行羽毛護理,確保羽毛處於最佳狀態,以應對長途飛行的挑戰。良好的羽毛狀態不僅能夠提高飛行效率,還能夠在惡劣天氣條件下提供更好的保護。
科研人員通過觀察鳥類的羽毛護理行為,可以獲得許多有價值的資訊。例如,羽毛護理行為的頻率和強度可能反映了個體的健康狀況和壓力水準。異常的羽毛護理行為可能是疾病或環境問題的早期指標。此外,通過研究不同物種的羽毛護理策略,科學家們可以更好地理解鳥類的生態適應和進化過程。
羽毛護理還與鳥類的色素沉積和羽毛結構維護有關。某些鳥類物種的羽毛顏色部分依賴於特定的微結構,這些結構需要通過精心的護理來維持。例如,藍色羽毛的顏色通常由羽毛中的微結構產生的光散射效應形成,而不是色素。因此,保持這些微結構的完整性對於維持羽毛的顏色至關重要。
最後,值得注意的是,羽毛護理行為是鳥類與環境互動的重要方式之一。通過羽毛護理,鳥類不僅維護自身的健康和功能,還能夠適應不同的生態環境和生活方式。這種適應性的行為模式充分展示了鳥類在進化過程中所形成的精巧生存策略,也為我們理解生物與環境的相互作用提供了寶貴的見解。
第五章鳥類飛行
5.1鳥類如何飛行
鳥類飛行是自然界中最令人驚嘆的現象之一,這種能力是經過數百萬年進化而來的複雜適應結果。鳥類飛行的關鍵在於其獨特的身體結構和生理特徵,這些特徵使得鳥類能夠克服重力,在空中自如地移動。
首先,鳥類的骨骼系統經過特殊改造,既輕巧又堅固。鳥類的骨骼內部呈蜂窩狀結構,充滿氣囊,大大減輕了身體重量。同時,某些骨骼如胸骨特別發達,為強大的飛行肌肉提供了附著點。鳥類的翅膀實際上是由前肢演化而來,其中的骨骼結構經過特化,能夠支撐飛行所需的羽毛。
羽毛是鳥類飛行的另一個關鍵要素。飛行羽毛(Flightfeathers)特別是初級飛羽和次級飛羽,形成了翅膀的主要升力面。這些羽毛的結構精密,能夠在飛行中調整形狀,以適應不同的飛行需求。羽毛不僅輕盈,還具有極高的強度,能夠承受飛行過程中的巨大應力。
鳥類的肌肉系統也經過特殊適應。飛行肌肉,尤其是胸大肌,佔鳥類體重的很大比例,為飛行提供了強大的動力。這些肌肉不僅力量大,還具有特殊的生理特性,能夠長時間持續工作而不疲勞。
鳥類的呼吸系統是另一個飛行適應的奇蹟。鳥類擁有獨特的氣囊系統,這種系統不僅提高了呼吸效率,還能在飛行中持續為肌肉提供充足的氧氣。氣囊系統還有助於調節體溫,這在高強度飛行中尤為重要。
鳥類的心血管系統也經過特殊改造,以支援高強度的飛行活動。鳥類的心臟相對體型而言更大,跳動更快,能夠高效地將氧氣和營養輸送到全身,特別是飛行肌肉。
在飛行力學方面,鳥類通過翅膀的拍打產生升力和推力。翅膀的上下拍動不僅推動鳥類向前,還通過改變翅膀的角度和形狀產生升力。不同的鳥類物種根據其生態位和飛行需求,evolve出不同的翅膀形狀和飛行模式。
鳥類的飛行控制系統也非常精密。尾羽在轉向和平衡中起著關鍵作用,而翅膀上的飛羽可以單獨調整,以實現精確的飛行控制。鳥類的大腦中有專門的區域負責處理飛行相關的資訊,使得鳥類能夠在複雜的三維空間中精確導航。
人類對鳥類飛行的研究不僅加深了我們對生物進化的理解,還為航空技術的發展提供了寶貴的靈感。通過研究鳥類飛行,科學家們正在開發更高效、更靈活的飛行器,這些技術可能在未來徹底改變我們的交通和運輸方式。
5.2鳥類飛行的空氣動力學
鳥類飛行的空氣動力學是一個極其複雜而fascinating的領域,它涉及了物理學、生物學和工程學的多個方面。鳥類通過數百萬年的進化,已經發展出了極其高效和靈活的飛行能力,這種能力的核心就在於其獨特的空氣動力學特性。
首先,我們需要理解飛行的基本原理。飛行主要依賴於四個力的平衡:升力、重力、推力和阻力。升力是使鳥類能夠克服重力保持在空中的關鍵力量。在鳥類飛行中,升力主要由翅膀產生。鳥類的翅膀橫截面呈現出特殊的形狀,稱為翼型(Airfoil)。這種形狀使得空氣在翅膀上下表面的流動速度不同,根據伯努利原理,這種速度差異會在翅膀上下表面之間產生壓力差,從而產生向上的升力。
鳥類翅膀的結構極其精妙。翅膀由堅韌而彈性的骨骼支撐,覆蓋著形狀特殊的飛羽。這些飛羽不僅輕盈,還具有驚人的強度和彈性。在飛行過程中,飛羽能夠根據需要改變形狀,這種動態調整能力大大提高了飛行的效率和靈活性。例如,在上升氣流中,鳥類可以展開翅膀,增大翼面積以獲得最大升力;而在俯衝時,則可以收縮翅膀以減少阻力。
鳥類飛行的一個關鍵特點是其翅膀的拍動。與固定翼飛行器不同,鳥類通過不斷拍打翅膀來產生升力和推力。這種拍動過程極其複雜,涉及翅膀的上下運動以及前後扭轉。在下拍過程中,翅膀以一定角度向下後方拍打,同時產生升力和向前的推力。在上拍過程中,翅膀則會稍微折疊並調整角度,以減少負升力。這種複雜的運動模式使得鳥類能夠在飛行中保持穩定,同時具備極高的機動性。
鳥類飛行中的另一個重要因素是尾羽的作用。尾羽不僅用於平衡和穩定,還在轉向和制動中起著關鍵作用。通過調整尾羽的展開程度和角度,鳥類可以精確控制其飛行方向和速度。在某些情況下,尾羽甚至可以作為額外的升力面,幫助鳥類在低速飛行或懸停時保持平衡。
鳥類飛行的效率很大程度上取決於其翅膀的形狀和大小。不同的鳥類物種因其生態位和飛行需求的不同,evolve出了各種不同的翅膀形態。例如,海鷗等滑翔鳥類通常擁有細長的翅膀,這種翅膀形狀能夠產生強大的升力,適合長時間的滑翔飛行。相比之下,鷹隼等獵食性鳥類則擁有寬闊而圓潤的翅膀,這種翅膀形狀有利於快速加速和靈活轉向,適合在林間追逐獵物。
鳥類飛行中的一個重要概念是翼展比(Aspectratio),即翅膀長度與寬度之比。高翼展比的翅膀(如信天翁)能夠產生更大的升力,適合長距離飛行;而低翼展比的翅膀(如麻雀)則有利於快速起飛和靈活轉向。鳥類通過調整翼展比來平衡飛行效率和機動性的需求。
在飛行過程中,鳥類還需要克服空氣阻力。為了減少阻力,鳥類evolve出了流線型的身體結構。它們的頭部和頸部能夠在飛行中收縮,形成與身體一體的流線型外形。此外,鳥類的羽毛表面具有微觀結構,能夠減少空氣摩擦,進一步降低阻力。
鳥類飛行中的一個有趣現像是所謂的「尾流利用」(Wakeutilization)。在群體飛行中,後方的鳥類可以利用前方鳥類翅膀產生的上升氣流,從而減少自身的能量消耗。這就是我們常見的大雁「V」字形隊列飛行的原理。通過這種方式,整個群體可以顯著提高飛行效率,尤其是在長距離遷徙時。
鳥類飛行的另一個重要方面是它們對氣流的利用能力。鳥類能夠敏銳地感知周圍的氣流變化,並迅速調整其飛行姿態以適應。例如,它們可以利用上升氣流(如熱氣流)來增加高度,而無需消耗太多能量。在山區飛行時,鳥類還能夠利用山坡產生的上升氣流來維持高度,這種技巧被稱為「坡面滑翔」(Ridgesoaring)。
在低速飛行和懸停時,鳥類面臨著特殊的空氣動力學挑戰。某些鳥類,如蜂鳥,已經evolve出了特殊的飛行技巧來應對這些情況。蜂鳥能夠通過高頻率的「8」字形翅膀運動來產生足夠的升力,從而實現穩定的懸停。這種飛行方式在空氣動力學上極為複雜,涉及了非穩態流體力學的原理。
鳥類飛行的起飛和著陸階段也值得特別關注。在起飛時,鳥類需要克服較大的重力和地面效應。它們通常會通過快速奔跑或跳躍來獲得初始速度,同時大幅度拍打翅膀以產生足夠的升力。著陸時,鳥類則需要精確控制速度和姿態。它們通過展開翅膀和尾羽來增加空氣阻力,同時調整身體角度以平穩降落。某些水鳥在著陸時還會利用水面的阻力來輔助減速。
鳥類飛行的空氣動力學研究不僅對生物學有重要意義,還為航空工程提供了寶貴的靈感。例如,某些現代飛機的機翼設計就借鑒了鳥類翅膀的形態。可變形機翼技術的發展也受到了鳥類動態調整翅膀形狀的啟發。此外,鳥類飛行中的能量效率和靈活性也為開發新型無人機提供了重要參考。
鳥類飛行的空氣動力學原理還涉及了許多微觀層面的巧妙設計。例如,鳥類羽毛的微觀結構有助於控制邊界層流動,減少紊流,從而降低阻力。一些研究還發現,某些鳥類的飛羽邊緣具有鋸齒狀結構,這種結構可以減少飛行時產生的噪音,同時可能還有改善空氣動力學性能的作用。
鳥類飛行的空氣動力學特性還與其生理結構密切相關。例如,鳥類的肺部和氣囊系統不僅提高了呼吸效率,還在飛行中起到了「氣動控制」的作用。通過調節體內氣囊的充氣狀態,鳥類可以微調其重心位置和身體剛性,從而優化飛行性能。
5.3鳥類飛行的動力
鳥類飛行的動力是一個複雜而精妙的系統,涉及多個生理和解剖學方面的適應。這種動力系統是鳥類在漫長的進化過程中形成的,使它們能夠克服重力,在空中自由翱翔。
鳥類飛行的主要動力來源是強大的胸肌。鳥類的胸肌,特別是胸大肌(Pectoralismajor),佔體重的很大比例,有些飛行能力強的鳥類的胸肌甚至可以佔到體重的30%以上。這些肌肉附著在特化的胸骨上,胸骨上有一個突出的骨脊,稱為龍骨(Keel),為這些強大的肌肉提供了附著點。胸大肌負責翅膀的下拍動作,這是產生升力和推力的主要來源。
與胸大肌配合的是胸小肌(Pectoralisminor),它負責翅膀的上舉動作。雖然上舉動作不直接產生升力,但它對於完成完整的飛行循環至關重要。這兩組肌肉的協調工作使得鳥類能夠持續不斷地拍打翅膀,維持飛行。
鳥類的肌肉纖維也經過特殊適應。飛行肌肉主要由快速收縮的肌纖維組成,這些纖維能夠快速產生力量,適應高頻率的翅膀拍動。此外,這些肌肉還具有較高的有氧代謝能力,能夠長時間持續工作而不疲勞,這對於長途飛行尤為重要。
為了支持這種高強度的肌肉活動,鳥類的心血管系統也經過了特殊改造。鳥類的心臟相對體型而言更大,心率也更快。這使得心臟能夠快速而有效地將氧氣和營養輸送到飛行肌肉。鳥類的血液中紅細胞的比例也較高,提高了攜氧能力。
鳥類的呼吸系統是另一個支持飛行的關鍵適應。鳥類擁有獨特的氣囊系統,這種系統不僅提高了呼吸效率,還能在飛行中持續為肌肉提供充足的氧氣。氣囊系統還有助於調節體溫,這在高強度飛行中尤為重要。
能量供應是維持飛行的另一個重要方面。鳥類通過高效的消化系統和代謝過程來滿足飛行的能量需求。它們能夠快速消化和吸收食物,並將能量儲存在肌肉和肝臟中的肝糖原形式。一些遷徙鳥類甚至能夠在飛行前大量積累脂肪,作為長途飛行的能量來源。
鳥類的骨骼系統也為飛行提供了支援。鳥類的骨骼既輕巧又堅固,內部呈蜂窩狀結構,充滿氣囊,大大減輕了身體重量。同時,某些骨骼如胸骨特別發達,為強大的飛行肌肉提供了穩固的附著點。
鳥類的神經系統也經過特殊適應以支持飛行。它們的大腦中有專門的區域負責控制飛行動作和平衡,使得鳥類能夠在複雜的三維空間中精確導航和機動。此外,鳥類還擁有快速的反射能力,能夠迅速應對飛行中的各種情況。
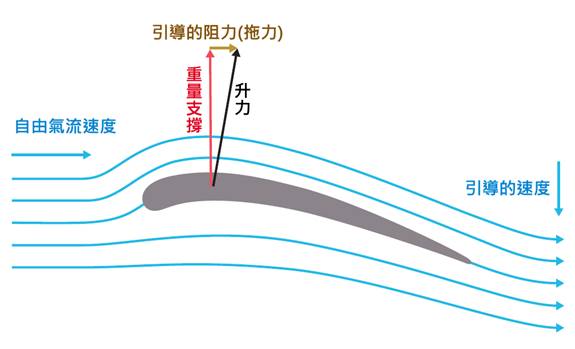
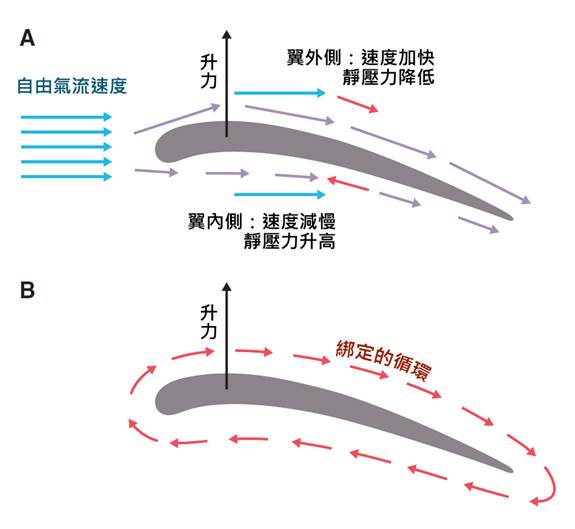
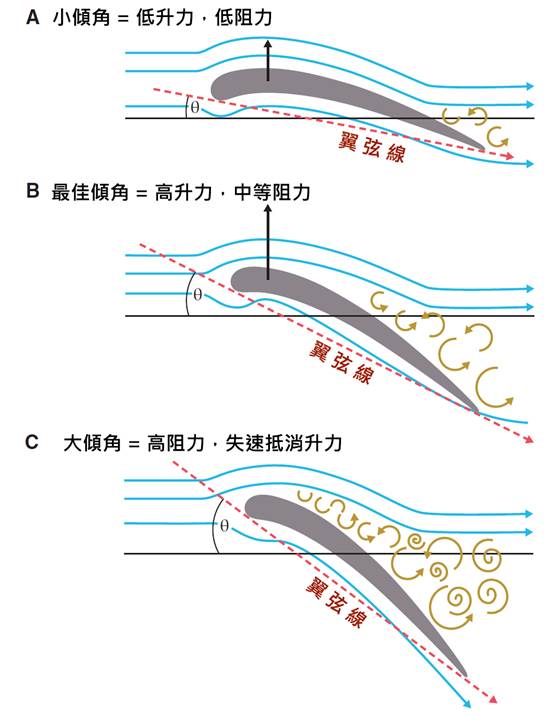

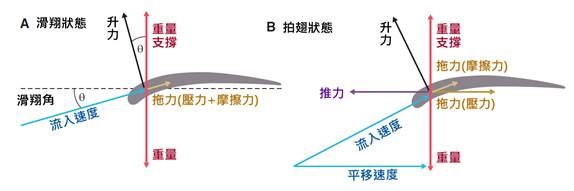
5.4鳥類拍翅飛行力學
鳥類的拍翅飛行力學是一個極其複雜而fascinating的領域,涉及了空氣動力學、生物力學和運動學的多個方面。拍翅飛行是鳥類最常見的飛行方式,它使鳥類能夠在空中保持穩定,同時具備極高的機動性和適應性。理解拍翅飛行力學不僅有助於我們更好地認識鳥類的生物學特性,還為航空工程和仿生技術的發展提供了重要的靈感來源。
拍翅飛行的基本原理是通過週期性的翅膀運動來產生升力和推力。在一個完整的拍翅週期中,鳥類的翅膀經歷下拍和上拍兩個主要階段。下拍階段是產生主要升力和推力的關鍵時期。在這個階段,翅膀以一定的角度向下後方拍打,空氣被向下和後方推動,根據牛頓第三定律,翅膀受到向上和向前的反作用力。這個力的向上分量就是升力,而向前分量則是推力。
上拍階段的力學特性則較為複雜。為了避免產生過多的負升力,鳥類會在上拍過程中稍微折疊翅膀,並調整翅膀的攻角。這種動作減少了上拍時的阻力,同時也能夠產生一定的正升力。某些鳥類,尤其是較小型的鳥類,甚至能夠在上拍階段產生顯著的升力,這種技巧被稱為「活躍上拍」(Activeupstroke)。
翅膀在拍動過程中不僅僅是簡單的上下運動,還涉及複雜的扭轉和彎曲。翅膀的這種動態變形能夠優化每個飛行階段的空氣動力學特性。例如,在下拍初期,翅膀會稍微向上扭轉,以獲得最佳的攻角;而在下拍後期,翅膀又會向下扭轉,以減少阻力。這種精細的控制要歸功於鳥類特殊的翅膀結構和強大的肌肉系統。
鳥類翅膀的結構對於拍翅飛行至關重要。翅膀由多個關節連接,這使得鳥類能夠靈活地改變翅膀的形狀和麵積。例如,在巡航飛行時,鳥類會部分折疊翅膀以減少翼展,從而降低能量消耗;而在需要快速加速或轉向時,則會完全展開翅膀以獲得最大的升力和控制力。此外,翅膀上的飛羽也能夠單獨調整,這種精細的控制能力使得鳥類能夠在不同的飛行階段優化其空氣動力學性能。
拍翅頻率是另一個影響飛行效率和性能的重要因素。不同大小和飛行風格的鳥類有著不同的最佳拍翅頻率。一般來說,體型較小的鳥類拍翅頻率較高,可以達到每秒數十次;而大型鳥類的拍翅頻率則相對較低,可能只有每秒幾次。拍翅頻率的選擇是一個權衡的結果,需要考慮能量消耗、飛行速度和機動性等多個因素。
鳥類拍翅飛行的一個重要特點是其非穩態空氣動力學特性。與固定翼飛行器不同,鳥類的翅膀在不斷運動中產生複雜的渦流結構。這些渦流可以被鳥類巧妙地利用來增加升力和推力。例如,在下拍結束時形成的前緣渦流可以在隨後的上拍階段被利用來產生額外的升力。這種動態失速(Dynamicstall)現象使得鳥類能夠在較大的攻角下飛行,從而獲得比穩態空氣動力學預測更高的升力系數。
拍翅飛行的效率很大程度上取決於翅膀的形狀和大小。不同的鳥類物種因其生態位和飛行需求的不同,evolve出了各種不同的翅膀形態。例如,燕子等敏捷飛行者通常擁有細長的翅膀,這種翅膀形狀有利於快速加速和靈活轉向。相比之下,信天翁等長距離飛行者則擁有細長而窄的翅膀,這種形狀能夠產生較大的升阻比,適合長時間的滑翔飛行。
在拍翅飛行過程中,鳥類還需要精確控制身體的姿態。尾羽在這個過程中起著關鍵作用。通過調整尾羽的展開程度和角度,鳥類可以改變身體的攻角,從而控制升力的大小和方向。此外,尾羽還在轉向和平衡中發揮重要作用,尤其是在低速飛行和懸停時。
鳥類的拍翅飛行還涉及了複雜的能量管理策略。為了提高飛行效率,許多鳥類採用間歇性飛行模式,即交替使用拍翅飛行和滑翔。這種飛行方式可以讓鳥類在滑翔階段休息肌肉,從而降低整體的能量消耗。某些鳥類,如啄木鳥,甚至發展出了波浪形飛行路徑,通過在上升階段積累勢能,然後在下降階段滑翔來節省能量。
拍翅飛行的力學特性在不同的飛行階段有所不同。起飛時,鳥類需要產生較大的升力來克服重力,因此通常會採用大幅度、高頻率的拍翅動作。在巡航飛行階段,拍翅動作會變得更加平緩和有節奏,以平衡升力、推力和能量消耗。著陸時,鳥類則需要通過調整翅膀和尾羽的位置來增加阻力,同時保持足夠的升力以控制下降速度。
鳥類的拍翅飛行還展現了驚人的適應性和靈活性。在面對不同的氣流條件時,鳥類能夠迅速調整其拍翅模式。例如,在遇到上升氣流時,鳥類可能會減少拍翅頻率,更多地依賴滑翔來節省能量。而在面對強逆風時,鳥類則可能會增加拍翅頻率和幅度以維持前進速度。
鳥類拍翅飛行的研究對於開發新型飛行器具有重要意義。例如,微型飛行器(Microairvehicles,MAVs)的設計就大量借鑒了鳥類和昆蟲的拍翅飛行原理。這些飛行器通過模仿生物的拍翅運動來產生升力和推力,具有高度的機動性和適應性,特別適合在複雜環境中執行任務。
此外,鳥類拍翅飛行的研究還涉及了許多前沿的科學問題。例如,鳥類如何在複雜的三維空間中精確控制其飛行路徑?它們如何感知和利用周圍的氣流資訊?這些問題的研究不僅有助於我們更好地理解鳥類的生物學特性,還可能為開發更高效、更智慧的飛行系統提供靈感。
拍翅飛行力學的研究方法也在不斷進步。高速攝影技術使我們能夠詳細觀察鳥類翅膀的運動軌跡;計算流體動力學(CFD)模擬則幫助我們理解翅膀周圍複雜的氣流結構;而先進的運動捕捉系統和力學感測器使我們能夠精確測量飛行過程中的各種物理參數。這些技術的發展極大地推動了我們對拍翅飛行力學的理解。
5.5鳥類滑翔與翱翔
滑翔與翱翔是鳥類飛行中極為重要且令人著迷的兩種模式,它們展現了鳥類對空中環境的高度適應能力和對能量利用的巧妙策略。雖然這兩個術語有時會被交替使用,但在嚴格的定義中,它們代表了不同的飛行技巧。
滑翔(Gliding)指的是鳥類在不拍動翅膀的情況下,利用重力和初始動能在空中前進的飛行方式。在滑翔過程中,鳥類通過調整翅膀的形狀和角度來控制下降速度和飛行方向。滑翔是一種相對簡單但高效的飛行方式,能夠讓鳥類在消耗極少能量的情況下覆蓋相當長的距離。
翱翔(Soaring)則是一種更為高級的飛行技巧,指的是鳥類利用上升氣流來獲得高度或維持高度的飛行方式。在翱翔過程中,鳥類幾乎不需要拍動翅膀,而是巧妙地利用環境中的氣流來提供升力。翱翔可以分為幾種主要類型,包括熱氣流翱翔、坡面翱翔和動態翱翔。
熱氣流翱翔是最常見的翱翔形式之一。當地面被太陽加熱時,空氣會上升形成熱氣流。鳥類能夠感知這些上升氣流,並在其中盤旋上升。一旦達到足夠的高度,鳥類就可以滑翔到下一個熱氣流。這種技巧被禿鷹、鷹類和鸛類等大型鳥類廣泛使用,使它們能夠在幾乎不消耗能量的情況下長時間飛行並覆蓋大distances。
坡面翱翔則是利用地形產生的上升氣流。當風吹到山坡或懸崖時,會被迫向上流動,形成持續的上升氣流。海鷗和信天翁等海鳥經常使用這種技巧在海岸線附近飛行。同樣,一些陸地鳥類也會利用山脈或丘陵產生的上升氣流進行長距離飛行。
動態翱翔是一種特殊的翱翔技巧,主要被一些海鳥如信天翁使用。這種技巧利用了海面上方風速隨高度增加的現象。鳥類通過在不同高度之間循環飛行,利用風速差來獲得升力和動能。這種飛行方式使得信天翁能夠在幾乎不消耗能量的情況下跨越vast的海洋距離。
滑翔和翱翔的效率很大程度上取決於鳥類的翅膀形態。長而窄的翅膀,如信天翁的翅膀,具有很高的升阻比,非常適合長時間的滑翔和翱翔。這種翅膀形狀能夠產生強大的升力,同時minimizing阻力,使鳥類能夠以極低的能量消耗維持飛行。
另一個影響滑翔和翱翔效率的重要因素是翼載荷,即鳥類的體重與翅膀面積之比。較低的翼載荷通常意味著更好的滑翔性能,因為鳥類可以在較低的速度下產生足夠的升力。然而,這也可能意味著在強風條件下的控制能力較差。因此,不同的鳥類物種根據其生態位和飛行需求evolve出了不同的翼載荷。
在滑翔和翱翔過程中,鳥類展現出了驚人的控制能力。通過微調翅膀的形狀、角度和相對於身體的位置,鳥類能夠精確地控制其飛行路徑和速度。尾羽在這個過程中也起著重要作用,幫助鳥類保持平衡並進行精細的方向控制。
一些鳥類物種還evolve出了特殊的適應性來提高滑翔和翱翔的效率。例如,某些禿鷹物種的翅膀末端羽毛可以分開,形成「指狀」結構。這種結構能夠減少翼尖渦流,從而降低誘導阻力,提高飛行效率。同樣,一些海鳥的翅膀能夠「鎖定」在展開位置,這使得它們能夠長時間保持翱翔而不會感到疲勞。
滑翔和翱翔技巧在鳥類的遷徙中起著至關重要的作用。許多長距離遷徙的鳥類物種大量依賴這些技巧來節省能量。例如,白鸛在從歐洲到非洲的遷徙過程中,大部分時間都在利用熱氣流翱翔。通過這種方式,它們能夠在幾乎不消耗體內儲備的情況下跨越撒哈拉沙漠這樣的巨大障礙。
滑翔和翱翔的能力還影響了鳥類的覓食策略。例如,某些猛禽物種會利用翱翔來搜尋獵物,這使得它們能夠在minimal能量消耗的情況下覆蓋大面積的區域。同樣,一些海鳥物種利用動態翱翔在vast的海洋面積上搜尋食物。
值得注意的是,並非所有鳥類都擅長滑翔和翱翔。這些技巧通常與鳥類的生態位和生存策略密切相關。例如,森林中的小型鳥類可能更依賴快速、靈活的拍翅飛行,而不是長時間的滑翔或翱翔。
滑翔和翱翔的研究對於航空工程和生物仿生學有重要意義。通過研究鳥類如何有效地利用氣流和minimizing能量消耗,科學家和工程師們可以開發更高效的飛行器。例如,無人機技術就借鑒了鳥類的滑翔和翱翔技巧,開發出能夠長時間在空中停留的太陽能飛行器。
此外,對滑翔和翱翔的研究還有助於我們理解大氣動力學和氣候變化。鳥類的飛行行為可以作為大氣條件的指標,幫助氣象學家更好地理解和預測天氣模式。例如,通過觀察鳥類的翱翔行為,我們可以獲得關於熱氣流分佈和強度的valuable信息。
滑翔和翱翔技巧的演化過程也是一個fascinating的研究領域。這些複雜的飛行技巧是如何在進化過程中逐步完善的?它們與鳥類的其他適應性特徵(如視覺系統、神經系統)之間有何聯繫?這些問題的研究不僅有助於我們理解鳥類的演化歷史,還可能為我們理解複雜行為的進化提供insights。
最後,值得一提的是,滑翔和翱翔技巧的研究還涉及了許多跨學科的問題。例如,鳥類如何感知和利用複雜的大氣條件?它們的大腦如何處理飛行中的各種資訊並做出快速決策?這些問題的研究需要結合氣象學、神經科學、行為學等多個學科的知識,體現了生物學研究的跨學科。
5.6鳥類能量效率與飛行速度
鳥類的能量效率和飛行速度是鳥類學和航空生物學中極為重要且fascinating的研究領域。這兩個方面不僅反映了鳥類的生理特性和適應能力,還直接影響著它們的生存策略和生態行為。理解鳥類如何平衡能量消耗和飛行速度,對於我們認識鳥類的生態學地位、遷徙模式以及進化過程都具有重要意義。
首先,我們需要認識到鳥類飛行的能量效率是一個複雜的概念,涉及多個因素的相互作用。鳥類在飛行過程中需要克服重力和空氣阻力,同時還要產生足夠的升力和推力。這些力的平衡需要消耗大量能量,而鳥類通過各種適應性策略來優化能量使用。
鳥類的飛行能量效率通常用每單位距離消耗的能量來衡量。這個指標受到多個因素的影響,包括鳥類的體型、翅膀形態、飛行方式以及環境條件等。一般來說,較大型的鳥類在長距離飛行中往往表現出更高的能量效率。這是因為它們的體表面積與體積之比較小,減少了熱量損失,同時它們通常具有更高的升阻比,能夠更有效地利用滑翔和翱翔技巧。
翅膀的形態對飛行能量效率有顯著影響。長而窄的翅膀,如信天翁的翅膀,具有很高的升阻比,非常適合長時間的滑翔和翱翔。這種翅膀形狀能夠在minimal能量消耗的情況下產生強大的升力。相比之下,短而寬的翅膀雖然在機動性方面更有優勢,但在長距離飛行中的能量效率較低。
飛行方式也是影響能量效率的關鍵因素。拍翅飛行雖然能提供極大的機動性,但能量消耗較高。相比之下,滑翔和翱翔能夠大幅降低能量消耗。許多長距離遷徙的鳥類物種採用間歇性飛行策略,即交替使用拍翅飛行和滑翔,以優化能量使用。例如,候鳥在遷徙過程中常常利用熱氣流上升到高空,然後長距離滑翔,這種策略能夠顯著降低整體能量消耗。
環境因素,如風速和方向、氣溫、海拔等,也會顯著影響飛行的能量效率。順風飛行可以大幅降低能量消耗,而逆風飛行則會增加能量需求。鳥類通常會選擇有利的氣象條件進行長距離飛行,有時甚至會等待合適的風向才開始遷徙。
轉向飛行速度,這是另一個與能量效率密切相關的重要參數。鳥類的飛行速度範圍很廣,從蜂鳥的懸停飛行到雨燕的高速飛行,展現了驚人的多樣性。飛行速度的選擇是多重因素權衡的結果,包括能量消耗、遷徙需求、捕食或避敵等。
每種鳥類都有其最佳巡航速度,即單位元距離能量消耗最低的速度。這個速度通常比理論上的最小功率速度略高。有趣的是,許多鳥類在長距離飛行時會選擇比最佳巡航速度稍快的速度飛行。這種策略雖然會略微增加能量消耗,但能夠更快地到達目的地,可能在整體上更有利於生存。
鳥類的最大飛行速度受到多個因素的限制,包括肌肉力量、心血管系統的能力、呼吸系統的效率等。一些鳥類物種能夠在短時間內達到驚人的速度,例如遊隼在俯衝時速度可超過300公里/小時,是已知最快的鳥類。然而,這種高速飛行通常只能維持很短的時間,能量消耗極大。
飛行速度還與鳥類的生態位和生存策略密切相關。例如,捕食性鳥類通常需要較高的速度和加速能力來捕獲獵物,而一些陸地鳥類可能更注重敏捷性和快速起飛能力來躲避捕食者。遷徙鳥類則需要在速度和耐力之間找到平衡,以完成長距離飛行。
值得注意的是,鳥類的飛行速度和能量效率會隨著飛行距離的變化而變化。在短距離飛行中,鳥類可能會選擇較高的速度,因為總能量消耗仍然相對較低。而在長距離飛行中,它們可能會選擇更接近最佳巡航速度的速度,以優化能量使用。
鳥類還evolve出了一些特殊的適應性來提高飛行的能量效率。例如,一些物種能夠在飛行過程中進入單側大腦睡眠狀態,這使得它們能夠在長時間飛行中得到休息。另外,一些鳥類在長距離飛行前會顯著增加體內脂肪儲備,為飛行提供充足的能量來源。
群體飛行是另一種提高能量效率的策略。許多鳥類物種在長距離飛行時會採用「V」字形隊列飛行。這種隊形能夠利用前方鳥類產生的上升氣流,從而減少後方鳥類的能量消耗。研究表明,這種策略可以使整個群體的能量效率提高約70%。
鳥類的飛行性能還受到季節性變化的影響。例如,在繁殖季節,許多鳥類會增加體重以儲存能量,這可能會降低它們的飛行速度和機動性。相反,在遷徙季節,鳥類通常會減輕體重以提高飛行效率。這種季節性的適應展示了鳥類在不同生命階段如何權衡飛行性能和其他生理需求。
研究鳥類的飛行能量效率和速度對於多個領域都有重要意義。在生態學方面,這些研究有助於我們理解鳥類的分佈範圍、遷徙路線選擇以及對氣候變化的響應。在進化生物學方面,它們幫助我們理解鳥類飛行能力的演化過程以及不同飛行策略的適應性意義。
在工程學領域,鳥類飛行的研究為開發高效的飛行器提供了寶貴的insights。例如,無人機技術的發展就大量借鑒了鳥類的飛行原理,特別是在提高能量效率和飛行持久性方面。同樣,對鳥類飛行速度的研究也為高速飛行器的設計提供了靈感。
研究方法的進步極大地推動了我們對鳥類飛行能量效率和速度的理解。GPS追蹤技術使我們能夠詳細記錄鳥類的飛行路徑和速度;高速攝影和計算機模擬則幫助我們分析飛行過程中的細節;而代謝率測量技術則使我們能夠準確評估飛行過程中的能量消耗。
然而,儘管我們在理解鳥類飛行方面取得了巨大進展,但仍有許多問題有待解答。例如,鳥類如何在複雜多變的環境中實時優化其飛行策略?它們的神經系統如何處理和整合各種飛行相關的信息?這些問題的解答可能需要更深入的跨學科研究,結合生物學、物理學、工程學等多個領域的知識。
5.7鳥類飛行能量學
飛行能量學是鳥類學研究中一個極為重要的領域,它探討鳥類在飛行過程中的能量消耗和利用情況。這個領域的研究不僅對理解鳥類的生理生態學特性至關重要,還為我們認識鳥類的行為模式、遷徙策略和進化適應提供了重要的視角。
鳥類飛行是一項高能耗活動,需要大量的能量來產生升力和推力,以克服重力和空氣阻力。飛行能量的來源主要是鳥類體內儲存的脂肪和糖原,這些物質在飛行肌肉中被氧化分解,釋放能量。鳥類的呼吸系統和循環系統經過特殊適應,能夠高效地為飛行肌肉提供氧氣和營養,並迅速清除代謝產物。
飛行能量消耗受多種因素影響,包括鳥類的體型、飛行速度、飛行方式以及環境條件等。一般來說,較大體型的鳥類在單位時間內的能量消耗較高,但如果以每公斤體重計算,大型鳥類的能量效率反而更高。這是因為它們的體表面積與體積比較小,減少了熱量損失,同時它們通常具有更高的升阻比,能夠更有效地利用滑翔和翱翔技巧。
飛行速度對能量消耗有顯著影響。每種鳥類都有其最佳巡航速度,即單位元距離能量消耗最低的速度。這個速度通常比理論上的最小功率速度略高。有趣的是,許多鳥類在長距離飛行時會選擇比最佳巡航速度稍快的速度飛行。這種策略雖然會略微增加能量消耗,但能夠更快地到達目的地,可能在整體上更有利於生存。
飛行方式也是影響能量消耗的重要因素。拍翅飛行雖然能提供極大的機動性,但能量消耗較高。相比之下,滑翔和翱翔能夠大幅降低能量消耗。許多長距離遷徙的鳥類採用間歇性飛行策略,即交替使用拍翅飛行和滑翔,以優化能量使用。例如,候鳥在遷徙過程中常常利用熱氣流上升到高空,然後長距離滑翔,這種策略能夠顯著降低整體能量消耗。
環境因素,如風速和方向、氣溫、海拔等,也會顯著影響飛行的能量消耗。順風飛行可以大幅降低能量消耗,而逆風飛行則會增加能量需求。鳥類通常會選擇有利的氣象條件進行長距離飛行,有時甚至會等待合適的風向才開始遷徙。
群體飛行是另一種降低能量消耗的策略。許多鳥類在長距離飛行時會採用「V」字形隊列飛行。這種隊形能夠利用前方鳥類產生的上升氣流,從而減少後方鳥類的能量消耗。研究表明,這種策略可以使整個群體的能量效率提高約70%。
5.8鳥類飛行機動性與穩定性
鳥類的飛行能力一直是科學家和工程師研究的重點,其中機動性和穩定性是兩個關鍵因素。鳥類通過長期進化,在這兩方面都達到了令人驚嘆的平衡。
鳥類的機動性主要體現在其靈活的翅膀結構和強大的胸肌。翅膀由多個關節組成,能夠在飛行中迅速改變形狀和角度。這種adaptability讓鳥類能夠在空中快速轉向、加速或減速。尤其是小型鳥類,如蜂鳥,更是機動性的典範,能夠在空中懸停甚至倒飛。大型猛禽如老鷹則能夠利用氣流滑翔長距離,同時保持敏捷的轉向能力以捕捉獵物。
穩定性方面,鳥類的身體結構也經過了精心的設計。首先,鳥類的重心位於翅膀下方,這種配置提供了自然的穩定性。其次,尾羽在飛行中起著類似飛機方向舵的作用,通過調整尾羽的形狀和角度,鳥類可以維持飛行的穩定性。此外,鳥類的骨骼結構輕盈中空,減輕了整體重量,同時保持了必要的強度。
鳥類的神經系統和感官也在飛行的穩定性中發揮重要作用。它們擁有敏銳的視力和平衡感,能夠迅速感知周圍環境的變化並做出相應調整。例如,在強風中飛行時,鳥類能夠不斷微調翅膀和尾部的位置以保持穩定。
有趣的是,不同種類的鳥類在機動性和穩定性之間取得了不同的平衡。比如,獵鷹注重高速和敏捷性,而albatross則更專注於長距離穩定飛行。這種多樣性反映了鳥類對不同生態環境的適應。
鳥類的飛行特性一直是人類航空技術的靈感來源。現代飛機設計中,許多概念都來自對鳥類飛行的觀察和研究。例如,一些飛機採用了可變幾何翼設計,模仿鳥類調整翅膀形狀的能力。無人機技術更是大量借鑒了小型鳥類的飛行特性,開發出了能夠高度機動的飛行器。
研究鳥類的飛行不僅對航空領域有重要意義,也幫助我們更好地理解和保護這些奇妙的生物。隨著技術的進步,科學家們能夠更精確地測量和分析鳥類的飛行數據,這些資訊對於生態保護和生物多樣性研究都具有重要價值。
鳥類飛行的機動性和穩定性是自然界的奇跡,體現了進化過程中的精妙平衡。通過對這一領域的持續研究,人類不僅能夠改進航空技術,還能增進對自然世界的理解和欣賞。鳥類的飛行能力提醒我們,自然界中蘊含著無窮的智慧和啟示,值得我們不斷探索和學習。
5.9鳥類飛行控制與尾部功能
鳥類的飛行控制和尾部功能是一個複雜而迷人的研究領域,涉及了生物學、空氣動力學和進化學等多個學科。這兩個方面在鳥類的飛行中扮演著至關重要的角色,共同構成了鳥類獨特而高效的飛行系統。
飛行控制是鳥類能夠在空中自如移動的關鍵。鳥類通過精細控制翅膀、尾部和身體其他部位來實現複雜的飛行動作。翅膀是最主要的控制結構,鳥類可以通過改變翅膀的形狀、角度和拍動頻率來調整升力和推力。例如,在需要快速加速時,鳥類會增加翅膀拍動的頻率和幅度;而在滑翔時,則會將翅膀展開以最大化升力。
鳥類的翅膀結構非常複雜,包含多個關節和肌肉群,使得它們能夠進行精細的動作控制。Primaryfeathers(主飛羽)位於翅膀的前緣和尖端,主要用於產生推力和升力。Secondaryfeathers(次飛羽)位於翅膀的後緣,幫助調整氣流並增加升力。Alula(翼擴)是位於翅膀前緣的一小組羽毛,在低速飛行時特別重要,可以防止失速並提高機動性。
鳥類的神經系統在飛行控制中起著核心作用。它們擁有高度發達的小腦,負責協調複雜的運動。視覺系統提供了關於周圍環境的即時資訊,而前庭系統則負責感知身體的方位和平衡。這些感官系統與運動控制系統緊密結合,使鳥類能夠在飛行中快速做出反應和調整。
尾部在鳥類飛行控制中的作用同樣不可忽視。尾部主要由尾羽組成,可以展開、收縮、上下擺動和左右轉動。這種多功能性使得尾部成為飛行控制的重要工具。尾部的主要功能包括:
1.穩定性控制:尾部可以像飛機的水準尾翼一樣,通過上下擺動來調整俯仰角,維持飛行的穩定性。在突然的氣流變化中,尾部的快速調整可以幫助鳥類保持平衡。
2.轉向輔助:通過左右擺動或不對稱展開,尾部可以協助鳥類進行轉向。這在低速飛行或滑翔時特別有效。
3.減速和著陸:當鳥類需要減速或準備著陸時,會將尾部完全展開,增加空氣阻力。這種動作類似於飛機降落時放下減速板。
4.升力產生:在某些飛行階段,特別是低速飛行時,展開的尾部可以產生額外的升力,幫助鳥類保持高度或進行精確的懸停。
5.信號傳遞:雖然不直接與飛行控制相關,但許多鳥類物種使用尾部展示來進行交流,這在求偶和領地防衛中尤為常見。
不同種類的鳥類因其生活環境和飛行需求的不同,演化出了各具特色的尾部結構。例如,燕子的分叉尾有助於高速飛行中的靈活轉向;孔雀的大型裝飾性尾羽雖然在飛行中可能不太實用,但在求偶展示中發揮重要作用;而蜂鳥的小巧尾部則有助於它們在空中保持精確的位置控制。
鳥類的飛行控制系統還包括一些特殊的適應性結構。例如,許多鳥類物種具有Airsacs(氣囊),這些與肺部相連的薄壁囊不僅提高了呼吸效率,還可以通過改變體內氣體分佈來微調重心位置,從而影響飛行姿態。另外,一些鳥類的飛羽可以分開,形成"翼槽",這種結構可以減少翅尖渦流,提高飛行效率。
研究鳥類的飛行控制和尾部功能不僅對於理解生物進化具有重要意義,也為人類航空技術的發展提供了寶貴的靈感。例如,一些現代飛機的機翼設計就借鑒了鳥類翅膀的可變形特性,以適應不同的飛行階段。無人機技術更是大量參考了鳥類的飛行控制機制,特別是在開發小型、高度機動的飛行器方面。
在生物保護領域,深入理解鳥類的飛行機制也有重要應用。例如,通過研究候鳥的長距離飛行策略,科學家們可以更好地預測和保護它們的遷徙路線。此外,這些研究還有助於我們評估人類活動對鳥類飛行能力的潛在影響,如城市化對鳥類飛行路線的改變等。
隨著科技的進步,研究鳥類飛行的方法也在不斷創新。高速攝影技術使得科學家能夠捕捉到鳥類飛行中的微小細節;風洞實驗則幫助研究者模擬不同飛行條件下鳥類的表現;而計算機模擬技術更是讓我們能夠在虛擬環境中測試各種飛行理論。這些先進的研究方法正在幫助我們揭示更多鳥類飛行的奧秘。
鳥類的飛行控制和尾部功能是一個極其複雜且精妙的系統,反映了自然界中進化的神奇力量。這個系統不僅保證了鳥類在空中的生存能力,也展現了生物適應性的極限。通過持續深入的研究,我們不僅能夠更好地理解和欣賞這些天空中的精靈,還能將這些知識應用到技術創新和環境保護中,為人類社會的發展做出貢獻。
鳥類飛行的奧秘仍有許多待解之處,每一項新的發現都可能為我們開啟新的視角,幫助我們更深入地理解生命的多樣性和自然界的奧妙。因此,對鳥類飛行控制和尾部功能的研究將繼續是一個充滿活力和前景的領域,吸引著來自不同學科背景的科學家共同探索。這些研究不僅豐富了我們對自然界的認知,也不斷啟發著人類在航空、機器人技術等領域的創新。通過跨學科的合作和持續的探索,我們有望在未來揭示更多關於鳥類飛行的秘密,並將這些知識轉化為實際應用,造福人類社會和自然環境。
5.10鳥類懸停、起飛與著陸
鳥類的飛行能力一直是自然界中最令人驚嘆的現象之一,其中懸停、起飛和著陸這三個飛行階段尤其引人注目。這些動作不僅展現了鳥類驚人的飛行技巧,也反映了它們對不同環境的適應能力。
懸停是鳥類飛行中最為精妙的技能之一。雖然並非所有鳥類都能夠懸停,但那些掌握這項技能的鳥類展現了令人驚嘆的控制力和精確度。蜂鳥是懸停的大師,它們能夠在空中保持幾乎完全靜止的狀態,這種能力使它們能夠有效地從花朵中吸取花蜜。蜂鳥懸停的關鍵在於其獨特的翅膀結構和運動方式。它們的翅膀能夠以驚人的速度(每秒可達50次以上)做"8"字形運動,在向上和向下擺動時都能產生升力。這種高效的翅膀運動使蜂鳥能夠在空中保持穩定位置,甚至能夠向後飛行。
除了蜂鳥,一些猛禽如鷹和隼也能夠短暫懸停,尤其是在搜尋獵物時。它們的懸停技術與蜂鳥不同,通常是通過快速拍打翅膀並調整身體角度來抵消重力。這種懸停雖然不如蜂鳥持久,但足以讓它們在空中保持穩定以便觀察地面。
水鳥如翠鳥也展示了特殊的懸停能力。它們能夠在水面上方保持相對靜止的位置,這種能力對於捕捉水中的魚類至關重要。翠鳥通過快速拍打翅膀並微調尾部位置來實現這種精確的懸停。
起飛是鳥類飛行中的關鍵階段,不同種類的鳥類有著不同的起飛策略。大多數小型鳥類,如麻雀或雀鷹,能夠從靜止狀態直接躍入空中。這種起飛方式需要強大的腿部肌肉和翅膀,鳥類會先用腿部發力跳躍,同時快速拍打翅膀產生初始升力。這個過程中,鳥類會迅速將翅膀完全展開,以最大化升力面積。
水鳥如天鵝或鵝在水面起飛時,會先在水面滑行一段距離,同時快速拍打翅膀。這種"跑道"式起飛幫助它們逐漸建立足夠的速度和升力。相比之下,海鳥如信天翁通常需要借助強風或高處起飛,因為它們的翅膀更適合滑翔而非快速拍打。
大型鳥類如鶴或鴕鳥,由於體重較大,起飛時需要更長的"跑道"。它們會先加速奔跑,同時逐漸展開翅膀,直到獲得足夠的速度和升力才能離地。這種起飛方式類似於飛機的起飛過程。
一些特殊的鳥類如茶隼,能夠利用上升氣流快速獲得高度。它們會尋找熱氣流,然後以螺旋式上升,這種技巧使它們能夠在很短的時間內達到很高的高度。
著陸是鳥類飛行中最具挑戰性的階段之一,需要精確的時間控制和空間感知。大多數鳥類在著陸時會採用類似的基本策略:減速、降低高度、展開翅膀和尾部以增加空氣阻力,最後伸出腿部準備接觸地面。
小型鳥類如麻雀或金絲雀通常能夠快速而靈活地著陸。它們會在接近目標時迅速減速,幾乎垂直下降,在最後一刻展開翅膀和尾部以緩衝衝擊。這種著陸方式允許它們在較小的空間內精確著陸,如細小的樹枝上。
大型鳥類如鶴或鵜鶘則需要更長的減速距離。它們通常會在空中盤旋以降低高度和速度,然後採取較淺的進場角度。在接近地面時,這些鳥類會將腿部伸出,同時將翅膀和尾部完全展開,形成類似降落傘的效果以進一步減速。
水鳥如鴨子或天鵝在水面著陸時會採用滑行技術。它們會將腳向前伸出,輕觸水面,同時保持翅膀部分展開以維持平衡。這種技術允許它們在水面上滑行一段距離,逐漸減速直到完全停止。
一些特殊的鳥類如海鳥中的信天翁,由於其翅膀結構主要適合長距離滑翔,在著陸時往往顯得不那麼優雅。它們可能需要多次嘗試才能成功著陸,特別是在風力較大的情況下。
鳥類的懸停、起飛和著陸能力是長期進化的結果,反映了它們對特定生態環境的適應。這些能力不僅幫助鳥類有效地捕食、逃避捕食者和遷徙,也使它們能夠利用各種不同的棲息地。
研究鳥類的這些飛行階段不僅對生物學和生態學有重要意義,也為人類在航空和機器人技術領域提供了寶貴的靈感。例如,垂直起降飛機的設計就借鑒了鳥類的懸停和垂直起飛能力。無人機技術也大量參考了鳥類的飛行機制,特別是在開發小型、高度機動的飛行器方面。
此外,瞭解鳥類的飛行特性對於野生動物保護也具有重要意義。例如,通過研究候鳥的起飛和著陸行為,科學家們可以更好地設計和保護它們的棲息地和中途停留站。在城市規劃中,這些知識也可以幫助減少建築物對鳥類飛行的幹擾,降低鳥類撞擊建築物的風險。
隨著科技的進步,研究鳥類飛行的方法也在不斷創新。高速攝影和3D動作捕捉技術使得科學家能夠精確記錄和分析鳥類飛行的每一個細節。風洞實驗和計算流體動力學模擬則幫助研究者深入理解鳥類飛行中的空氣動力學原理。這些先進的研究方法正在幫助我們揭示更多關於鳥類飛行的奧秘。
鳥類的懸停、起飛和著陸能力展示了自然界的奇妙和生物進化的力量。這些看似簡單的動作背後,蘊含著複雜的生物力學原理和精妙的控制機制。通過持續深入的研究,我們不僅能夠更好地理解和欣賞這些天空中的精靈,還能將這些知識應用到技術創新和環境保護中,推動人類社會的可持續發展。
鳥類飛行的研究仍有很長的路要走。每一項新的發現都可能為我們開啟新的視角,幫助我們更深入地理解生命的多樣性和自然界的奧妙。因此,對鳥類飛行,特別是懸停、起飛和著陸這些關鍵階段的研究將繼續是一個充滿活力和前景的領域,吸引著來自不同學科背景的科學家共同探索。這些研究不僅豐富了我們對自然界的認知,也不斷啟發著人類在航空、機器人技術等領域的創新。通過跨學科的合作和持續的探索,我們有望在未來揭示更多關於鳥類飛行的秘密,並將這些知識轉化為實際應用,造福人類社會和自然環境。
5.11鳥類飛行適應與特化
鳥類的飛行適應與特化是進化過程中的一個奇蹟,展現了自然選擇的力量和生物多樣性的豐富。這些適應和特化涵蓋了鳥類的解剖結構、生理機能和行為模式,使它們能夠在各種環境中高效飛行。
首先,鳥類的骨骼系統經過了特殊的進化。它們的骨骼既輕盈又堅固,許多骨頭是中空的,filledwithairsacs(充滿氣囊),這大大減輕了整體重量而不失強度。胸骨特別發達,形成了竭狀突起(keel),為強大的飛行肌肉提供了附著點。翅膀骨骼也經過改造,形成了適合飛行的扇形結構。
鳥類的羽毛是飛行適應中最顯著的特徵之一。飛羽結構複雜,輕盈而堅韌,能夠在提供足夠升力的同時保持彈性。不同部位的羽毛具有不同功能,例如初級飛羽(primaryfeathers)主要用於產生推力,而次級飛羽(secondaryfeathers)則幫助產生升力。羽毛還具有保溫和防水功能,對飛行至關重要。
鳥類的呼吸系統也經過了特殊適應。它們擁有高效的氣囊系統,能夠在吸氣和呼氣時都進行氣體交換,這種雙向流動的呼吸方式大大提高了氧氣利用率,為高強度飛行提供了充足的能量支援。
在生理方面,鳥類的心臟相對體型來說更大更強壯,能夠支援飛行時的高代謝需求。它們的血液中紅血球數量也較高,提高了攜氧能力。某些鳥類,特別是遷徙鳥,還能夠在飛行前大量儲存脂肪作為長途飛行的能量來源。
不同的飛行特化反映了鳥類對特定環境的適應。例如,信天翁擁有細長的翅膀,適合長時間滑翔;蜂鳥的翅膀相對較小,能夠快速拍打,適合懸停和靈活飛行;而禿鷹的寬大翅膀則適合利用熱氣流盤旋。
某些鳥類還發展出了特殊的飛行技巧。比如,一些海鳥能夠利用海面上的風層進行動態滑翔(dynamicsoaring),這種技術使它們能夠在幾乎不消耗能量的情況下飛行長距離。另一方面,像燕子這樣的鳥類則發展出了高速飛行和靈活轉向的能力,有助於捕捉空中的昆蟲。
鳥類的飛行適應與特化不僅體現在身體結構上,還反映在它們的行為和生活方式中。例如,許多鳥類會根據季節變化調整它們的飛行模式,有些甚至會進行長距離遷徙。這種行為適應使它們能夠利用不同地區的資源,並避開不利的氣候條件。
5.12翅膀類型與飛行方式
鳥類的翅膀類型和飛行方式是生物學和空氣動力學研究的重要領域,反映了鳥類對不同生態環境的適應和進化。這些特徵不僅決定了鳥類的飛行能力,還影響了它們的生存策略和生態位。
鳥類的翅膀類型可以大致分為幾種主要形態,每種形態都與特定的飛行方式和生態適應相關聯。高頻率翅膀(Highfrequencywings)是最常見的類型之一,通常見於小型鳥類如麻雀、finches(雀科鳥類)和warblers(鶯科鳥類)。這種翅膀相對較短而圓,適合在樹林或灌木叢中快速機動飛行。高頻率翅膀允許鳥類進行快速起飛和敏捷轉向,這對於捕食昆蟲或逃避捕食者至關重要。這類鳥類的飛行方式通常是快速拍打翅膀,能夠在狹小空間內靈活操作。
與之相對的是低頻率翅膀(Lowfrequencywings),常見於大型鳥類如鷺鷥、herons(蒼鷺)和storks(鸛)。這種翅膀通常較長且寬大,適合緩慢而持久的飛行。低頻率翅膀的鳥類通常採用滑翔和滑行的飛行方式,能夠利用上升氣流長時間保持在空中而無需頻繁拍打翅膀。這種飛行方式非常節能,適合長距離遷徙或在開闊地區尋找食物。
高速翅膀(Highspeedwings)是另一種特化的翅膀類型,常見於燕子、swifts(雨燕)和falcons(隼)等鳥類。這種翅膀細長而尖銳,形狀類似新月或鐮刀,非常適合高速飛行。高速翅膀的鳥類能夠達到驚人的飛行速度,有些物種甚至能夠在俯衝時達到每小時200公里以上。這種翅膀形態使得鳥類能夠快速捕捉飛行中的昆蟲或在空中追逐其他鳥類。
長距離飛行者如信天翁和albatrosses(信天翁)則擁有超長的細長翅膀,這種翅膀形態被稱為滑翔翅膀(Soaringwings)。滑翔翅膀的特點是翼展極大但翅膀寬度較窄,翼展與翅膀寬度的比例(aspectratio)非常高。這種設計使得鳥類能夠利用海洋上的風層進行動態滑翔(dynamicsoaring),幾乎不消耗能量就能飛行數千公里。滑翔翅膀的鳥類通常生活在開闊的海洋環境中,它們的飛行方式主要依賴於風力和空氣動力學原理,而不是頻繁拍打翅膀。
還有一種特殊的翅膀類型是短翼(Ellipticalwings),常見於森林鳥類如鴿子、woodpeckers(啄木鳥)和一些雉科鳥類。這種翅膀相對較短且寬圓,適合在密集的樹林中快速起飛和靈活飛行。短翼允許鳥類在狹小空間內進行快速而精確的機動,這對於在複雜環境中躲避障礙物或捕食者非常有利。
蜂鳥擁有一種獨特的翅膀類型,通常被稱為懸停翅膀(Hoveringwings)。這種翅膀相對較小但肌肉發達,能夠進行高頻率的"8"字形運動,使蜂鳥能夠在空中保持靜止或甚至向後飛行。懸停翅膀的設計使蜂鳥成為自然界中唯一能夠長時間懸停的鳥類,這種能力對於它們從花朵中吸取花蜜至關重要。
鳥類的飛行方式與其翅膀類型密切相關,但也受到其他因素的影響,如體型、生態位和行為特徵。例如,很多猛禽如鷹和鷂擁有寬大的翅膀,適合利用熱氣流進行盤旋和滑翔。這種飛行方式使它們能夠在空中長時間搜尋獵物,同時節省能量。當發現獵物時,這些鳥類又能迅速轉換為俯衝攻擊的飛行方式。
水鳥如鴨子和geese(鵝)通常具有相對較短而強壯的翅膀,適合快速起飛和在水面上滑行。這些鳥類的飛行方式通常是持續快速拍打翅膀,能夠在較短距離內迅速獲得高度。相比之下,像pelicans(鵜鶘)這樣的大型水鳥則擁有寬大的翅膀,更適合滑翔和利用氣流。
某些鳥類還發展出了特殊的飛行技巧。例如,禿鷹和其他大型猛禽能夠利用熱氣流進行螺旋上升(thermalsoaring),這種技術使它們能夠在很短的時間內達到很高的高度。另一方面,一些海鳥如shearwaters(海燕)能夠利用海面上的風層進行波浪滑翔(wavesoaring),這種飛行方式使它們能夠在長距離飛行中節省大量能量。
鳥類的翅膀類型和飛行方式還與它們的遷徙行為密切相關。長距離遷徙鳥類通常擁有能夠高效滑翔的翅膀,這使它們能夠在長途飛行中節省能量。例如,北極燕鷗擁有流線型的翅膀,能夠進行世界上最長的鳥類遷徙,每年往返南北極。
研究鳥類的翅膀類型和飛行方式不僅對生物學和生態學有重要意義,也為工程領域提供了寶貴的靈感。例如,無人機技術的發展就大量借鑒了不同鳥類的飛行特性。一些研究者正在開發能夠模仿鳥類翅膀變形的靈活機翼,以提高飛行器的效率和適應性。
此外,瞭解不同鳥類的飛行特性對於野生動物保護也具有重要意義。例如,在設計風力發電場時,考慮當地鳥類的飛行高度和方式可以幫助減少鳥類碰撞風機的風險。在城市規劃中,瞭解鳥類的飛行需求也可以幫助設計更加生態友好的建築和綠地系統。
隨著科技的進步,研究鳥類翅膀和飛行的方法也在不斷創新。高速攝影和計算機模擬技術使得科學家能夠更精確地分析鳥類飛行的空氣動力學特性。風洞實驗和野外追蹤技術則提供了更多關於鳥類飛行效率和行為的數據。這些先進的研究方法正在幫助我們揭示更多關於鳥類翅膀進化和飛行適應的奧秘。
鳥類的翅膀類型和飛行方式是自然選擇的傑作,展示了生物如何通過長期進化適應不同的生態環境。這個領域的研究不僅豐富了我們對自然界的理解,也為人類在航空、機器人和環境保護等領域的創新提供了無窮的靈感。通過持續深入的研究和跨學科的合作,我們有望在未來揭示更多關於鳥類飛行的秘密,並將這些知識轉化為實際應用,推動科技進步和生態保護。
5.13鳥類飛行環境適應
鳥類的環境適應是一個複雜而迷人的研究領域,體現了生物進化的奇妙過程。鳥類作為地球上分佈最廣的脊椎動物之一,已經適應了從極地到熱帶,從海洋到沙漠的各種環境。這種廣泛的適應性不僅體現在它們的身體結構上,還反映在行為、生理和生態特徵中。
在極地環境中,鳥類面臨著嚴寒和食物稀缺的挑戰。帝企鵝(Emperorpenguin)是極地適應的典範。它們擁有厚實的脂肪層和密集的羽毛,這些特徵能夠有效地阻隔寒冷。帝企鵝還進化出了特殊的社交行為,在嚴冬時期,它們會聚集在一起形成緊密的群體,輪流站在外圍以抵禦寒風。極地的其他鳥類如雪鴞(Snowyowl)則進化出了白色的羽毛,不僅能夠提供保暖,還能在雪地中進行偽裝。
相比之下,熱帶環境的鳥類需要應對高溫和濕度。許多熱帶鳥類如鸚鵡科(Psittacidae)的鳥類擁有鮮艷的羽毛,這不僅用於吸引配偶,還可能有助於反射過多的陽光。一些熱帶鳥類如禿鷲(Vulture)進化出了禿頭,這有助於體溫調節。在行為上,許多熱帶鳥類會在一天中最熱的時候躲在樹蔭下休息,只在早晚較涼爽的時候活動。
沙漠環境對鳥類提出了獨特的挑戰,主要是缺水和極端溫度。沙漠鳥類如鴕鳥(Ostrich)進化出了高效的水分保持機制。它們能夠忍受體溫的較大波動,並且有特殊的鼻腔結構可以減少呼吸時的水分損失。一些小型沙漠鳥類如沙百靈(Sandlark)則通過在白天躲在陰涼處,只在清晨和黃昏活動來適應高溫。
海洋環境的鳥類面臨著完全不同的挑戰。海鳥如信天翁(Albatross)擁有特殊的鹽腺,可以排出體內多餘的鹽分,這使它們能夠飲用海水。海鳥的羽毛也有特殊的結構,能夠防水並保持體溫。在行為上,許多海鳥如企鵝科(Spheniscidae)的成員進化出了出色的游泳和潛水能力,能夠在水下捕捉魚類。
森林環境的鳥類則發展出了適應樹冠生活的特徵。例如,巨嘴鳥(Toucan)的大型喙不僅用於取食果實,還有助於在茂密的樹冠中保持平衡。啄木鳥科(Picidae)的鳥類則進化出了特殊的頭骨結構和強壯的喙,能夠在樹幹上鑿洞築巢和尋找昆蟲。許多森林鳥類如鸚鵡還進化出了靈活的腳趾,能夠牢固地抓住樹枝。
高山環境對鳥類提出了低氧和低溫的挑戰。高山鳥類如安第斯禿鷲(Andeancondor)進化出了更大的肺部和更高的血紅蛋白含量,以適應低氧環境。一些高山鳥類如雪雞(Snowcock)則發展出了特殊的羽毛結構,能夠在極端寒冷的環境中保持體溫。
城市環境作為一種相對新的生態系統,也促使鳥類進行了快速適應。城市鳥類如鴿子(Pigeon)和麻雀(Sparrow)表現出了較強的環境適應能力。它們學會了利用人造結構築巢,適應了人類食物,甚至改變了自己的鳴叫頻率以克服城市噪音的幹擾。
鳥類的環境適應還體現在它們的遷徙行為上。許多鳥類通過季節性遷徙來適應環境的週期性變化。例如,北極燕鷗(Arctictern)每年往返南北極,這是地球上最長的動物遷徙路線。這種遷徙行為使得鳥類能夠利用不同地理位置的季節性資源,避開不利的氣候條件。
在生理適應方面,鳥類展現出了驚人的多樣性。例如,蜂鳥(Hummingbird)能夠將體溫降低到接近環境溫度的水準,進入一種類似冬眠的狀態(torpor),以度過寒冷的夜晚。而一些生活在高海拔地區的鳥類,如安第斯山脈的雀類,則進化出了特殊的血紅蛋白結構,能夠在低氧環境中更有效地運輸氧氣。
鳥類的環境適應還體現在它們的繁殖策略上。例如,在資源豐富但季節性強的溫帶地區,許多鳥類採取一年繁殖一次的策略,將繁殖時間安排在食物最豐富的季節。而在資源相對穩定的熱帶地區,一些鳥類可能全年都有繁殖活動。極地鳥類如帝企鵝則採取了極端的繁殖策略,在南極的嚴冬中孵化蛋並撫育幼鳥。
鳥類的環境適應還涉及到它們的飲食習慣。不同環境中的鳥類進化出了適應當地食物資源的喙部結構和消化系統。例如,達爾文雀(Darwin'sfinches)因其多樣化的喙部形態而聞名,這些不同形狀的喙適應了加拉帕戈斯群島上不同的食物資源。而食火烈鳥(Flamingo)則進化出了特殊的喙部結構和舌頭,能夠過濾水中的小型生物作為食物。
鳥類的環境適應研究不僅對生物學和生態學有重要意義,還為我們理解氣候變化對生物多樣性的影響提供了重要線索。通過研究鳥類如何適應不同環境,科學家們可以更好地預測和應對全球氣候變化可能帶來的生態影響。
此外,鳥類的環境適應研究還為生物工程和材料科學提供了靈感。例如,企鵝羽毛的防水結構已經被用於開發新型的防水材料,而蜂鳥的飛行機制則為開發微型飛行器提供了參考。
隨著科技的進步,研究鳥類環境適應的方法也在不斷創新。基因組學技術使得科學家能夠從分子水準理解鳥類的適應機制。衛星追蹤技術則使得研究者能夠詳細記錄鳥類的遷徙路線和行為模式。這些新技術正在幫助我們揭示更多關於鳥類環境適應的奧秘。
鳥類的環境適應是一個動態的過程,反映了生物與環境之間的複雜互動。通過研究鳥類如何適應不同的環境挑戰,我們不僅能夠更好地理解生物進化的過程,還能為生態保護和可持續發展提供重要的科學依據。這個領域的研究不僅豐富了我們對自然界的認識,也為人類在面對環境變化時提供了寶貴的借鑒。
5.14飛行中的導航與定向
鳥類在飛行中的導航和定向能力一直是科學界研究的熱點話題,這種能力不僅令人驚嘆,還為我們理解動物行為和地球物理學提供了寶貴的見解。鳥類能夠在沒有現代導航工具的情況下完成長距離遷徙,甚至能夠精確地返回特定的築巢地點,這種能力背後蘊含著複雜的生理機制和行為策略。
鳥類的導航系統可以被認為是一個多層次的綜合系統,包括視覺、嗅覺、聽覺,以及對地球磁場的感知等多種感官輸入。這些不同的感官資訊被鳥類的大腦整合,形成一個複雜而精確的導航地圖。
視覺導航是鳥類最基本也最重要的導航方式之一。鳥類擁有極其敏銳的視力,能夠識別遠處的地標和地形特徵。許多鳥類物種能夠記住遷徙路線上的關鍵地標,如山脈、河流或海岸線,並利用這些地標進行導航。一些研究表明,鳥類甚至可能利用星空進行夜間導航。例如,歐洲知更鳥(Europeanrobin)在實驗中表現出了使用星座圖進行方向定位的能力。
除了視覺,嗅覺也在鳥類導航中扮演著重要角色。一些研究表明,某些鳥類物種可能通過記憶和識別特定地區的氣味來輔助導航。這種能力在信鴿(Homingpigeon)身上表現得尤為明顯。實驗發現,當信鴿的嗅覺被阻斷時,它們返回家園的能力會受到顯著影響。科學家推測,鳥類可能通過建立一種"嗅覺地圖"來輔助長距離導航。
聽覺在鳥類導航中的作用相對較少被研究,但也不容忽視。一些鳥類物種可能利用低頻聲波來感知地形變化或遠處的海洋聲音。例如,有研究表明,一些海鳥可能利用海浪的聲音作為導航參考。
然而,鳥類最令人驚嘆的導航能力可能來自於它們感知地球磁場的能力。這種被稱為磁感應(Magnetoreception)的能力允許鳥類感知地球磁場的強度和傾角,從而確定自己的位置和方向。科學家們提出了幾種可能的磁感應機制,其中最受關注的是基於視網膜中特定蛋白質的量子機制和基於鐵磁物質的機制。
基於量子機制的理論認為,鳥類視網膜中的一種稱為隱花色素(Cryptochrome)的蛋白質可能對磁場敏感。當光子激發這種蛋白質時,會形成一對自由基,這對自由基的量子態可能受到地球磁場的影響。鳥類可能通過感知這種變化來獲取方向資訊。這個理論解釋了為什麼一些鳥類的磁感應能力似乎依賴於光線。
另一種理論則關注鳥類體內的鐵磁物質。科學家在鳥類的上喙和內耳中發現了含鐵的結構,這些結構可能像微小的羅盤一樣對地球磁場做出反應。這種機制可能允許鳥類直接感知磁場的強度和方向。
值得注意的是,鳥類可能同時使用多種導航機制,並根據不同的環境條件和飛行階段靈活切換。例如,在晴朗的日子裡,視覺導航可能起主導作用,而在陰天或夜間,磁感應可能變得更加重要。
鳥類的導航能力還表現在它們對時間的精確把握上。許多遷徙鳥類表現出驚人的時間準確性,能夠在幾乎相同的日期開始它們的季節性遷徙。這種能力被稱為生物鐘(Biologicalclock),它允許鳥類根據日光長度的變化來預測季節的變化,從而選擇最佳的遷徙時機。
鳥類的導航系統還表現出令人驚嘆的適應性和學習能力。年輕的鳥類通常需要跟隨成年個體學習遷徙路線,但一旦學會,它們就能夠獨立完成長距離飛行。一些研究甚至表明,某些鳥類物種能夠根據環境變化調整其遷徙路線,這種靈活性對於應對氣候變化和人類活動帶來的環境變化至關重要。
鳥類的導航能力還涉及到複雜的資訊處理和決策過程。在長距離飛行中,鳥類需要不斷評估自身的能量狀態、天氣條件和潛在的危險,並做出相應的調整。例如,候鳥在遇到不利的天氣條件時,可能會選擇改變飛行高度或暫時停留在中途站休息。這種動態決策能力是鳥類成功完成長距離遷徙的關鍵因素之一。
研究鳥類的導航和定向能力不僅有助於我們理解動物行為,還為人類技術發展提供了inspiration。例如,鳥類的磁感應能力已經inspire了新型導航技術的研發,這些技術可能在GPS信號不可用的情況下提供精確的定位。此外,研究鳥類如何在複雜的三維空間中導航,也為開發更先進的無人機控制系統提供了參考。
鳥類導航研究還對生態保護具有重要意義。隨著人類活動對自然環境的影響日益加劇,許多鳥類的遷徙路線和棲息地受到了威脅。理解鳥類的導航機制可以幫助我們更好地設計保護策略,例如保護關鍵的中途停留地或減少人造光源對夜間遷徙鳥類的幹擾。
隨著科技的進步,研究鳥類導航的方法也在不斷創新。衛星跟蹤技術使得科學家能夠精確記錄鳥類的遷徙路線和行為。神經生物學技術則允許研究者深入探索鳥類大腦中與導航相關的神經機制。基因組學研究也揭示了一些與導航能力相關的基因,為我們理解這種能力的進化起源提供了新的視角。
鳥類在飛行中的導航和定向能力是自然界中最令人驚嘆的現象之一,體現了生物進化的奇妙力量。這個領域的研究不僅豐富了我們對自然界的理解,還為解決人類面臨的技術挑戰提供了新的思路。通過持續深入的研究,我們有望揭示更多關於鳥類導航的奧秘,並將這些知識應用到更廣泛的領域中,推動科技進步和生態保護。
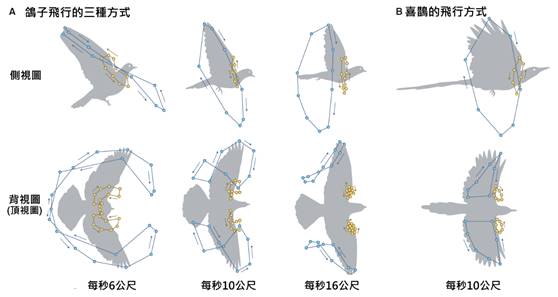
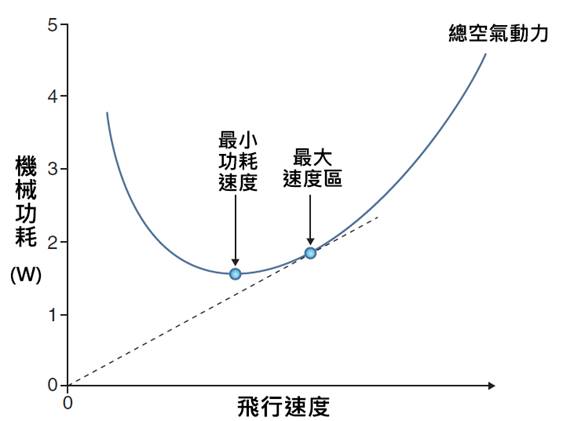
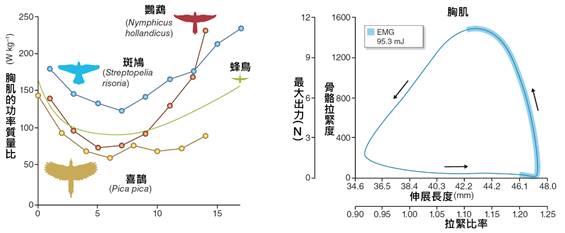
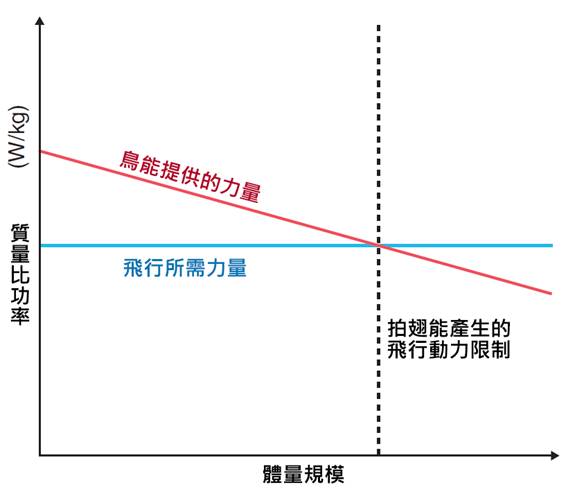
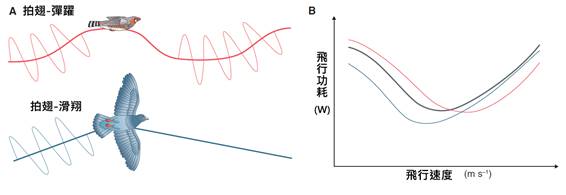
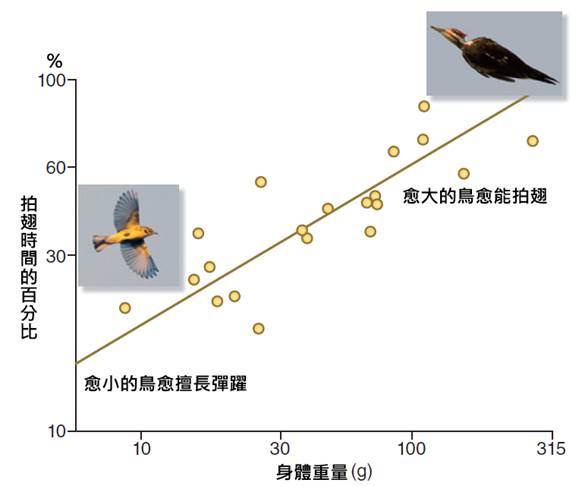
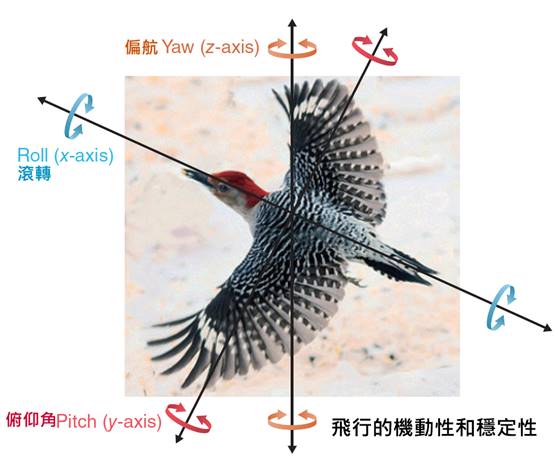


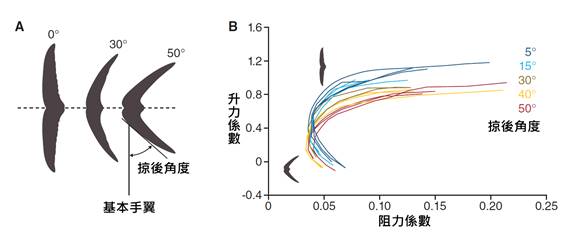
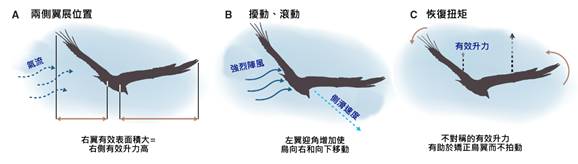
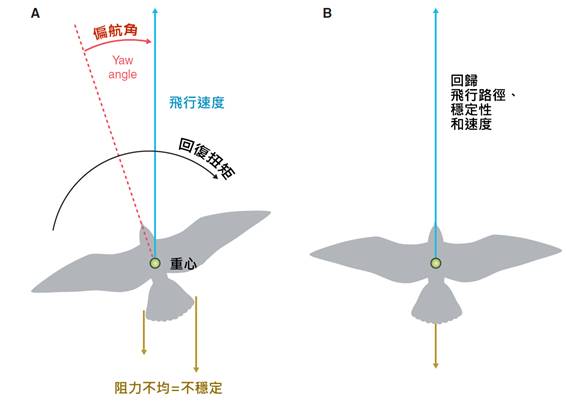
5.15能潛水鳥類水中運動
能夠潛水的鳥類及其水中運動是鳥類學和生物力學研究中一個極為引人入勝的領域。這些鳥類通過長期的進化,發展出了一系列獨特的適應性特徵,使它們能夠在水下環境中靈活移動和覓食。這種能力不僅展示了生物適應性的奇蹟,也為我們理解水下環境中的生存策略提供了寶貴的見解。
首先,我們需要認識到,能夠潛水的鳥類並不局限於某一個特定的分類群。事實上,潛水能力在鳥類中多次獨立進化,出現在不同的科屬中。其中最著名的潛水鳥類包括企鵝科(Spheniscidae)、鸊鷉科(Podicipedidae)、鷗鷺科(Alcidae)、鸕鶿科(Phalacrocoracidae)等。這些鳥類雖然在分類學上可能相距甚遠,但它們都表現出了驚人的水下運動能力。
企鵝可能是最為人熟知的潛水鳥類。它們的身體結構高度適應了水下生活,幾乎可以被視為飛行鳥類向水生生活方式轉變的極致例子。企鵝的翅膀已經進化成了類似鰭狀的結構,在水中能夠產生強大的推進力。它們的身體呈流線型,減少了水的阻力。企鵝的骨骼也比其他鳥類更加緻密,這減少了浮力,使它們能夠更容易地潛入水中。帝企鵝(Emperorpenguin)是潛水能力最強的鳥類之一,能夠潛到超過500米的深度,並能在水下停留長達20分鐘。
鸊鷉是另一類著名的潛水鳥類。它們雖然保留了飛行能力,但在水下的運動能力同樣出色。鸊鷉的腳掌位於身體後部,這種結構雖然使它們在陸地上行走不便,但在水中卻能提供強大的推進力。鸊鷉的羽毛結構也經過特殊適應,能夠調節浮力。通過調整體內氣囊的空氣量,鸊鷉能夠精確控制自己在水中的位置。
鷗鷺科的鳥類,如厚嘴海雀(Razorbill)和海雀(Commonmurre),展示了另一種適應水下生活的方式。這些鳥類保留了較好的飛行能力,同時也能夠在水下靈活運動。它們使用半張開的翅膀在水下"飛翔",這種運動方式既利用了翅膀的推進力,又保持了良好的機動性。
鸕鶿科的鳥類則以其出色的捕魚能力而聞名。這些鳥類能夠快速潛入水中,並在水下靈活地追逐魚群。鸕鶿的喙部結構特殊,能夠輕易抓住滑溜的魚類。它們的羽毛結構也經過特殊適應,能夠在潛水時減少浮力,而在離水後又能迅速恢復防水性能。
潛水鳥類的水下運動涉及多個生理和物理方面的適應。首先是呼吸系統的適應。這些鳥類通常具有較大的肺容量和更高效的氧氣利用率。一些物種還進化出了特殊的血紅蛋白結構,能夠在低氧環境中更有效地運輸氧氣。例如,帝企鵝的血紅蛋白對氧氣的親和力非常高,使它們能夠在深水中長時間活動。
潛水鳥類還需要應對水壓帶來的挑戰。深潛的鳥類,如某些企鵝物種,其骨骼和肌肉結構能夠承受極高的水壓。它們的氣囊系統也能夠在潛水時收縮,以避免壓力損傷。一些研究表明,潛水鳥類可能還具有特殊的神經系統適應,能夠在高壓環境下維持正常的神經功能。
在視覺方面,潛水鳥類也進行了特殊的適應。水下環境的光線條件與空氣中大不相同,潛水鳥類的眼睛結構經過調整以適應這種環境。例如,它們的角膜和晶狀體形狀經過改變,以補償水對光線折射的影響。一些深潛的物種,如帝企鵝,還進化出了對弱光敏感的視網膜結構,使它們能夠在深水中的微弱光線下捕獵。
潛水鳥類的體溫調節也是一個重要的適應方面。水的導熱性比空氣強得多,因此這些鳥類需要特殊的機制來維持體溫。它們通常具有厚實的脂肪層和高度發達的羽毛結構,能夠有效隔絕冷水。一些物種還進化出了特殊的血液循環系統,能夠在潛水時減少對四肢和外周組織的血液供應,將有限的熱量集中在維持核心器官的功能上。
潛水鳥類的運動方式也因物種而異。企鵝主要依靠翅膀(鰭)的擺動來推進,這種運動方式類似於海豚或海獅的游泳方式。鸊鷉和鷗鷺科的鳥類則主要使用腳蹼來推進,同時也可能輔以翅膀的動作。鸕鶿在追逐獵物時能夠展現出驚人的機動性,它們能夠快速改變方向,精確地跟蹤移動的魚群。
值得注意的是,許多潛水鳥類還保留了某種程度的飛行能力。這種雙重適應反映了自然選擇的平衡性,使這些鳥類能夠在水下和空中兩種環境中生存。然而,這種平衡通常意味著在某些方面的妥協。例如,能夠飛行的潛水鳥類通常不能潛入很深的水中,而最擅長深潛的企鵝則完全失去了飛行能力。
研究潛水鳥類的水中運動不僅對生物學有重要意義,也為工程學提供了寶貴的借鑒。例如,企鵝的流線型身體結構和游泳方式已經inspire了一些水下機器人的設計。研究這些鳥類如何在水下環境中有效導航和捕獵,也可能為開發新型的水下傳感器和導航系統提供靈感。
此外,瞭解潛水鳥類的行為和生理特性對於海洋生態系統的研究和保護也至關重要。這些鳥類往往是海洋生態系統健康狀況的重要指標。通過研究它們的潛水行為和覓食模式,科學家們可以獲得關於海洋環境變化和魚類種群動態的寶貴信息。
隨著技術的進步,研究潛水鳥類水中運動的方法也在不斷創新。微型攝像機和傳感器技術使得科學家能夠直接觀察和記錄這些鳥類在水下的行為。三維運動分析技術則幫助研究者精確量化潛水鳥類的運動模式。這些新技術正在幫助我們揭示更多關於潛水鳥類水中運動的奧秘。
潛水鳥類的水中運動能力是自然界適應性進化的一個絕佳例證。這些鳥類通過長期的演化,在形態、生理和行為上發展出了一系列獨特的適應特徵,使它們能夠在水下環境中靈活運動和高效覓食。研究這些適應性特徵不僅豐富了我們對生物多樣性的理解,也為解決人類在水下活動中面臨的技術挑戰提供了新的思路。
5.16水面捕魚鳥類的水中運動
水面捕魚鳥類的水中運動是一個極其引人入勝的研究領域,展示了鳥類對水生環境的獨特適應。這些鳥類雖然不像深潛的企鵝或鸊鷉那樣完全適應了水下生活,但它們在水面和淺水區域展現出了令人驚嘆的捕魚技巧和運動能力。這種特殊的生態位置使得水面捕魚鳥類成為了連接空中、水面和水下生態系統的重要紐帶。
在討論水面捕魚鳥類的水中運動之前,我們需要瞭解這個群體包括哪些主要的鳥類。常見的水面捕魚鳥類包括鷺科(Ardeidae)的鳥類如大白鷺(Greategret)和夜鷺(Nightheron),鸛科(Ciconiidae)的鳥類如白鸛(Whitestork),鷸科(Scolopacidae)的鳥類如濱鷸(Sandpiper),以及鷗科(Laridae)的鳥類如銀鷗(Herringgull)等。這些鳥類雖然在分類學上可能相距甚遠,但它們都進化出了在水面或淺水區域捕食的能力。
水面捕魚鳥類的水中運動主要可以分為幾種類型:涉水(Wading)、點水(Dipping)、浮游(Floating)和淺潛(Shallowdiving)。每種運動方式都與特定的形態特徵和捕食策略相關聯。
涉水是最常見的水中運動方式之一,尤其在鷺科和鸛科的鳥類中普遍存在。這些鳥類通常具有細長的腿,能夠在淺水區域緩慢而穩定地行走。例如,大白鷺在捕食時會小心翼翼地在水中移動,盡量減少水的擾動以避免驚動獵物。這種運動方式要求鳥類具有良好的平衡能力和精確的腳步控制。鷺鳥的腳趾通常較長且分開,增加了在軟泥底質上的穩定性。
點水是另一種常見的捕魚方式,特別是在鷗科和燕鷗科(Sternidae)的鳥類中。這種方法涉及鳥類在低空飛行,然後迅速俯衝並用喙或腳觸及水面捕捉魚類。銀鷗就經常使用這種方法捕捉靠近水面的魚類。這種運動方式需要鳥類具有精確的空間判斷能力和快速的反應速度。燕鷗科的鳥類如普通燕鷗(Commontern)更是點水捕魚的專家,它們能夠在高速飛行中精確地捕捉水面下的魚類。
浮游是一種相對靜態的水中運動方式,常見於鴨科(Anatidae)和部分鷗科的鳥類。這些鳥類能夠在水面上漂浮,並利用這個位置來觀察和捕捉周圍的獵物。例如,綠頭鴨(Mallard)常常在水面漂浮,同時用喙在水中過濾食物。這種運動方式要求鳥類具有良好的浮力控制能力。鴨科鳥類的羽毛結構特殊,能夠儲存空氣,提高浮力。同時,它們的腳掌呈蹼狀,可以在需要時提供推進力。
淺潛是一些水面捕魚鳥類使用的更為主動的捕食方法。例如,鸕鶿科(Phalacrocoracidae)的鳥類雖然主要以深潛捕魚為主,但也常在淺水區域進行短暫的潛水捕魚。這種運動方式要求鳥類能夠快速改變身體姿態,從水面迅速潛入水中。鸕鶿的羽毛結構特殊,在潛水時能夠吸水減少浮力,而在離水後又能迅速甩乾。
水面捕魚鳥類的這些水中運動方式都與其特定的形態特徵密切相關。例如,涉水鳥類通常具有細長的腿和喙,這有助於它們在淺水中行走和捕捉獵物。點水捕魚的鳥類則往往有強壯的翅膀和尖銳的喙,使它們能夠在快速飛行中精確捕獲獵物。浮游捕魚的鳥類通常身體較為圓潤,有利於在水面保持平衡。
這些鳥類在水中運動時還面臨著許多挑戰。首先是平衡問題。在不穩定的水面環境中保持平衡需要精細的肌肉控制和反應能力。其次是視覺適應。水面的反光和折射可能會影響鳥類對水下獵物的判斷,因此這些鳥類通常具有特殊的視覺適應,如能夠快速調整焦距的能力。
水溫也是一個重要因素。雖然這些鳥類不像深潛鳥類那樣長時間浸泡在水中,但它們仍然需要應對水溫帶來的熱量損失。許多水面捕魚鳥類都有發達的腿部血管網絡,可以有效調節體溫。同時,它們的羽毛結構也經過特殊適應,能夠在短時間接觸水後迅速恢復防水性能。
水面捕魚鳥類的捕食行為也反映了它們的水中運動能力。例如,鷺科鳥類在捕食時常常保持一種靜止的姿態,這需要極強的肌肉控制能力。當獵物出現時,它們能夠以閃電般的速度出擊,這種快速的運動要求鳥類具有出色的反應速度和精確的空間判斷能力。
一些水面捕魚鳥類還表現出了複雜的協作行為。例如,美洲鹮(Americanwhiteibis)有時會集體在淺水區域形成一條線,共同向前推進以驅趕魚類。這種行為不僅需要個體具備良好的水中運動能力,還要求群體之間有高度的協調性。
水面捕魚鳥類的研究對於理解水生生態系統的功能具有重要意義。這些鳥類往往是連接水陸生態系統的重要紐帶,它們的捕食行為可能對魚類種群產生顯著影響。同時,研究這些鳥類的水中運動也可能為水面和淺水區域的機器人設計提供inspiration。例如,鷺類鳥類的穩定步態可能為開發能夠在不穩定水面環境中運動的機器人提供參考。
隨著技術的進步,研究水面捕魚鳥類水中運動的方法也在不斷創新。高速攝影技術使得研究者能夠捕捉到這些鳥類快速運動的細節。水下攝像系統則提供了鳥類在水下短暫運動時的寶貴影像資料。此外,新型的生物遙測技術,如微型加速度計和陀螺儀,使得科學家能夠詳細記錄這些鳥類在自然環境中的運動模式。
水面捕魚鳥類的水中運動能力是自然選擇的產物,反映了生物對特定生態位的精細適應。這些鳥類通過長期進化,在形態、生理和行為上發展出了一系列獨特的特徵,使它們能夠在水陸交界處的複雜環境中有效捕食。研究這些適應性特徵不僅豐富了我們對生物多樣性的理解,也為解決人類在類似環境中面臨的技術挑戰提供了新的思路。通過深入研究水面捕魚鳥類的水中運動,我們可以更好地理解生態系統的複雜性,並為水生環境的保護和管理提供科學依據。
5.17水面遊弋鳥類的水域運動
水面遊弋鳥類的水域運動是鳥類學和生物力學研究中一個引人入勝的領域。這類鳥類雖然不像深潛鳥類那樣完全適應了水下生活,也不同於僅在水面捕魚的鳥類,而是在水面和淺水區域展現出了獨特的運動方式和生存策略。水面遊弋鳥類主要包括鴨科(Anatidae)、天鵝(Swans)、鵝(Geese)等水禽,以及一些鷗科(Laridae)和鸊鷉科(Podicipedidae)的成員。
這些鳥類的水域運動主要可以分為幾種類型:漂浮(Floating)、游泳(Swimming)、淺潛(Surfacediving)和濾食(Filterfeeding)。每種運動方式都與鳥類的特定形態特徵和生態位置密切相關。
漂浮是水面遊弋鳥類最基本的水域運動方式。這些鳥類能夠長時間在水面保持穩定的漂浮狀態,這得益於它們特殊的身體結構和羽毛特性。例如,鴨子和天鵝的身體呈流線型,有助於減少水的阻力。它們的羽毛具有特殊的結構,能夠儲存空氣,提高浮力。同時,這些鳥類能夠通過調整體內氣囊的空氣量來精確控制浮力。漂浮不僅是一種休息狀態,也是這些鳥類觀察周圍環境、尋找食物和避免捕食者的重要方式。
游泳是水面遊弋鳥類的另一個主要運動方式。這些鳥類通常具有蹼狀的腳掌,能夠在水中產生強大的推進力。游泳時,鳥類會有節奏地移動腳掌,同時保持身體在水面上的平衡。不同種類的水面遊弋鳥類有不同的游泳風格。例如,天鵝游泳時通常保持優雅的姿態,緩慢而穩定地前進。而一些鴨科鳥類,如綠頭鴨(Mallard),則能夠快速改變游泳方向和速度。
有趣的是,一些水面遊弋鳥類還發展出了一種特殊的游泳方式,稱為「漂滑」(Skimming)。這種方式通常見於一些鷗科的鳥類,如黑浮鷗(Blackskimmer)。它們會貼近水面快速飛行,同時將下喙浸入水中,藉此捕捉靠近水面的小魚或浮游生物。這種獨特的運動方式需要精確的平衡控制和敏銳的感知能力。
淺潛是一些水面遊弋鳥類採用的更為主動的捕食方法。例如,鸊鷉科的鳥類能夠從水面迅速潛入水中捕捉魚類。這種運動方式要求鳥類能夠快速改變身體姿態,從漂浮狀態轉換為潛水狀態。鸊鷉的羽毛結構特殊,能夠在潛水時吸水減少浮力,而在離水後又能迅速甩乾。一些鴨科鳥類,如綠頭鴨,也常常採用「倒立潛水」(Upending)的方式在淺水區域覓食,這種姿態要求鳥類具有良好的平衡能力和柔軟的頸部。
濾食是許多水面遊弋鳥類,特別是鴨科鳥類採用的一種特殊的覓食方式。這些鳥類的喙部內側有特殊的板狀結構,稱為「層疊板」(Lamellae)。當鳥類在水中游動時,會張開喙部,讓水流通過這些層疊板,從而過濾出水中的小型生物和植物碎屑。這種運動方式要求鳥類能夠精確控制頭部和頸部的運動,同時保持身體在水面上的穩定。
水面遊弋鳥類的這些水域運動方式都與其特定的形態特徵密切相關。例如,它們通常具有扁平的身體和寬大的胸骨,這有助於在水面保持穩定。它們的羽毛有特殊的結構,能夠防水並保持體溫。同時,這些鳥類的腿部通常位於身體後部,這雖然使它們在陸地上行走不便,但在水中卻能提供強大的推進力。
水面遊弋鳥類在水域運動時還面臨著多方面的挑戰。首先是體溫調節問題。水的導熱性比空氣強得多,因此這些鳥類需要特殊的機制來維持體溫。它們通常具有厚實的脂肪層和高度發達的羽毛結構,能夠有效隔絕冷水。同時,許多物種還進化出了特殊的血液循環系統,能夠減少對四肢的熱量損失。
其次是平衡問題。在不穩定的水面環境中保持平衡需要精細的肌肉控制和反應能力。水面遊弋鳥類通常具有發達的前庭系統,能夠精確感知身體的位置和姿態變化。此外,它們的尾部通常能夠靈活運動,起到類似船舵的作用,幫助調整方向和保持平衡。
視覺適應也是水面遊弋鳥類面臨的一個重要挑戰。水面的反光和折射可能會影響鳥類對水下環境的判斷。因此,這些鳥類通常具有特殊的視覺適應,如能夠快速調整焦距的能力,以及能夠同時看清水上和水下環境的雙焦視力。
水面遊弋鳥類的水域運動還涉及到複雜的能量平衡問題。游泳和濾食雖然消耗的能量相對較少,但長時間在冷水中活動仍然需要大量能量。因此,這些鳥類通常具有高效的消化系統和代謝機制,能夠快速吸收和利用食物中的能量。
一些水面遊弋鳥類還表現出了有趣的社交行為。例如,很多鴨科和天鵝科的鳥類會形成大型群體,在水面上共同活動。這種群體行為不僅有助於防禦捕食者,還可能提高覓食效率。群體中的個體需要具備良好的空間感知能力和社交協調能力,以避免在密集的群體中發生碰撞。
水面遊弋鳥類的研究對於理解水生生態系統的功能具有重要意義。這些鳥類往往是水生植物種子的重要傳播者,它們的活動可能對水生植被的分佈產生顯著影響。同時,它們的覓食行為也可能影響水體中的浮游生物和小型魚類種群。
此外,研究水面遊弋鳥類的水域運動對於水面船隻和機器人的設計也有啟發意義。例如,這些鳥類在水面保持穩定的能力可能為開發更穩定的小型水面設備提供參考。它們的濾食機制也可能inspire新型的水質淨化技術。
隨著技術的進步,研究水面遊弋鳥類水域運動的方法也在不斷創新。水下攝像系統提供了這些鳥類在水下短暫活動時的寶貴影像資料。新型的生物遙測技術,如微型加速度計和壓力感應器,使得科學家能夠詳細記錄這些鳥類在自然環境中的運動模式和能量消耗。此外,計算機模擬技術也被廣泛應用於研究這些鳥類的水動力學特性。
水面遊弋鳥類的水域運動能力是長期進化的結果,反映了生物對水陸交界生態位的精細適應。這些鳥類通過形態、生理和行為上的一系列適應,成功地在水面和淺水區域建立了自己的生態位置。研究它們的水域運動不僅能夠幫助我們更好地理解生物適應性的奧秘,也為解決人類在類似環境中面臨的技術挑戰提供了新的思路。通過深入研究水面遊弋鳥類的水域運動,我們可以更全面地認識水生生態系統的複雜性,為水生環境的保護和可持續利用提供科學依據。
5.18淺水灘地覓食鳥類的水域運動
淺水灘地覓食鳥類的水域運動是一個引人入勝的研究領域,展現了鳥類對特定環境的精細適應。這些鳥類主要活動在潮間帶、沼澤、泥灘等淺水區域,它們的水域運動方式既不同於深水潛水鳥類,也有別於水面遊弋鳥類,而是發展出了一套獨特的運動和覓食策略。
淺水灘地覓食鳥類主要包括鷸科(Scolopacidae)、鴴科(Charadriidae)、彩鷸科(Rostratulidae)、鷺科(Ardeidae)等多個科的成員。這些鳥類雖然在分類學上可能相距甚遠,但它們都適應了在淺水和泥灘環境中覓食的生活方式。
這些鳥類的水域運動主要可以分為幾種類型:涉水(Wading)、探測(Probing)、啄食(Pecking)和掃掠(Sweeping)。每種運動方式都與鳥類的特定形態特徵和生態位置密切相關。
涉水是淺水灘地覓食鳥類最基本的水域運動方式。這些鳥類通常具有細長的腿,能夠在淺水中穩定地行走。例如,大白鷺(Greategret)和黑頭髻鷸(Black-wingedstilt)就以其修長的腿部著稱,使它們能夠在不同深度的水中自如活動。涉水時,這些鳥類需要精確控制每一步的落點,以避免驚動獵物或陷入軟泥中。這種運動方式要求鳥類具有出色的平衡能力和精細的肌肉控制。
探測是許多涉禽,特別是鷸科鳥類常用的覓食方式。長嘴半蹼鷸(Long-billeddowitcher)和黑尾濱鷸(Bar-tailedgodwit)等物種擁有細長的喙,能夠深入泥土或沙中尋找藏匿的無脊椎動物。這種運動方式要求鳥類具有敏感的觸覺和靈活的頸部肌肉。有些物種,如歐洲厚嘴鷸(Eurasiancurlew),還能夠彎曲喙部以更好地探測彎曲的洞穴。
啄食是一種更為直接的覓食方式,常見於鴴科鳥類如金斑鴴(Europeangoldenplover)。這些鳥類通常有較短而堅固的喙,適合在水面或泥面上快速啄取小型獵物。啄食要求鳥類具有敏銳的視力和快速的反應能力。一些物種如反嘴鷸(Piedavocet)還發展出了特殊的側向啄食方式,能夠在淺水中掃過一定範圍尋找食物。
掃掠是一種特殊的覓食方式,主要見於一些特化的物種如鶴鷸(Eurasianspoonbill)。這些鳥類擁有扁平的喙,能夠在淺水中左右擺動,過濾出小型生物。這種運動方式要求鳥類能夠精確控制頭部運動,同時保持身體的穩定。
淺水灘地覓食鳥類的這些水域運動方式都與其特定的形態特徵密切相關。例如,它們通常具有細長的腿和特化的喙部結構。細長的腿不僅有助於在不同深度的水中活動,還能減少水的阻力。特化的喙部則適應了不同的覓食策略,從探測泥土到過濾水體,每種喙型都有其獨特的功能。
這些鳥類的腳部結構也經過了特殊的適應。有些物種如半蹼鷸(Semipalmatedsandpiper)的腳趾間有部分蹼,這增加了在軟泥上的支撐面積。而另一些物種如跳鷺(Greatjacana)則有極長的腳趾,能夠在浮葉植物上行走。這些適應使得不同物種能夠在各自的生態位置中高效活動和覓食。
淺水灘地覓食鳥類在水域運動時還面臨著多方面的挑戰。首先是平衡問題。在不穩定的淺水和泥灘環境中保持平衡需要精細的肌肉控制和反應能力。這些鳥類通常具有發達的前庭系統,能夠精確感知身體的位置和姿態變化。此外,許多物種還能夠靈活地使用翅膀輔助平衡,特別是在快速運動或遇到突發情況時。
其次是視覺適應。淺水環境中的光線條件複雜,水面的反光和折射可能會影響鳥類對水下獵物的判斷。因此,這些鳥類通常具有特殊的視覺適應,如能夠快速調整焦距的能力,以及在水上和水下環境中都能清晰視物的雙焦視力。有些物種如翻石鷸(Ruddyturnstone)還發展出了特殊的覓食行為,會翻動石頭和貝殼尋找藏匿的獵物,這需要精確的空間判斷能力。
觸覺在淺水灘地覓食鳥類的水域運動中也扮演著重要角色。許多物種的喙端有豐富的神經末梢,能夠感知微小的壓力變化。這使得它們能夠在視線不佳的泥土或濁水中準確定位獵物。例如,鷸鴕(Kiwi)雖然主要活動在陸地上,但其高度發達的觸覺系統與許多涉禽相似,展示了這種感官在覓食中的重要性。
能量平衡是淺水灘地覓食鳥類面臨的另一個挑戰。在淺水中不斷運動和覓食需要消耗大量能量,特別是在寒冷的環境中。因此,這些鳥類通常具有高效的消化系統和代謝機制。許多物種在遷徙前會積累大量脂肪,以應對長途飛行和潛在的食物短缺。同時,它們也發展出了靈活的覓食策略,能夠根據環境條件和食物可用性調整自己的行為。
潮汐變化對淺水灘地覓食鳥類的水域運動有顯著影響。這些鳥類需要根據潮汐節律調整自己的活動模式。例如,在漲潮時,一些物種會轉移到更高的地方或改變覓食策略。而在退潮時,它們則會跟隨水線移動,尋找新暴露的覓食區域。這種適應要求鳥類具有良好的時間感知能力和靈活的行為策略。
群體行為在淺水灘地覓食鳥類中也很常見。許多物種如黑腹濱鷸(Dunlin)常常形成大型群體共同覓食。這種行為不僅有助於防禦捕食者,還可能提高覓食效率。群體中的個體需要具備良好的空間感知能力和社交協調能力,以在密集的群體中有效活動而不互相幹擾。
研究淺水灘地覓食鳥類的水域運動對於理解濕地生態系統的功能具有重要意義。這些鳥類往往是連接不同營養級別的重要紐帶,它們的覓食活動可能對底棲無脊椎動物的種群動態產生顯著影響。同時,它們也是環境變化的敏感指標,通過監測這些鳥類的行為和種群變化,科學家們可以評估濕地生態系統的健康狀況。
此外,研究淺水灘地覓食鳥類的水域運動對於濕地保護和管理也有重要的實際意義。瞭解這些鳥類的活動模式和棲息地需求,可以幫助制定更有效的保護策略。例如,在設計人工濕地或恢復退化濕地時,可以根據目標物種的需求來優化水深、底質類型和植被結構。
隨著技術的進步,研究淺水灘地覓食鳥類水域運動的方法也在不斷創新。高速攝影技術使得研究者能夠捕捉到這些鳥類快速運動的細節。遙感技術和地理資訊系統(GIS)則幫助科學家們在更大的空間尺度上分析這些鳥類的活動模式。此外,新型的生物遙測技術,如微型GPS追蹤器和加速度計,使得科學家能夠詳細記錄這些鳥類在自然環境中的運動軌跡和能量消耗。
淺水灘地覓食鳥類的水域運動能力是長期進化的結果,反映了生物對特定生態位置的精細適應。這些鳥類通過形態、生理和行為上的一系列適應,成功地在充滿挑戰的濕地環境中建立了自己的生態位置。研究它們的水域運動不僅能夠幫助我們更好地理解生物適應性的奧秘,也為解決人類在類似環境中面臨的技術挑戰提供了新的思路。通過深入研究淺水灘地覓食鳥類的水域運動,我們可以更全面地認識濕地生態系統的複雜性,為濕地的保護和可持續利用提供科學依據。
5.19地棲鳥類的陸地運動
地棲鳥類的陸地運動是鳥類學和生物力學研究中一個獨特而引人入勝的領域。這類鳥類雖然保留了飛行能力,但主要在地面上活動,因此發展出了一系列適應陸地生活的特殊運動方式。地棲鳥類包括多個科的成員,如鴕鳥科(Struthionidae)、雉科(Phasianidae)、鶴科(Gruidae)、秧雞科(Rallidae)等,它們在形態、行為和生態位置上都表現出了對陸地生活的顯著適應。
地棲鳥類的陸地運動主要可以分為幾種類型:步行(Walking)、奔跑(Running)、跳躍(Jumping)和攀爬(Climbing)。每種運動方式都與鳥類的特定形態特徵和生態需求密切相關。
步行是地棲鳥類最基本的陸地運動方式。與飛行鳥類相比,地棲鳥類通常具有更強壯的腿部肌肉和更大的腳掌,這使它們能夠在地面上穩定地行走。例如,火雞(Turkey)和孔雀(Peafowl)就以其穩健的步態著稱。步行時,這些鳥類通常保持身體平衡,頭部會隨著每一步的節奏輕微擺動,這有助於穩定視線。一些大型地棲鳥類,如鴕鳥(Ostrich),甚至完全失去了飛行能力,但發展出了高效的雙足步行方式。
奔跑是許多地棲鳥類用於逃避捕食者或追逐獵物的重要運動方式。一些物種如鴕鳥和美洲鴕(Greaterrhea)能夠達到驚人的奔跑速度,鴕鳥的最高速度甚至可以超過每小時70公里。這種高速奔跑能力得益於它們強健的腿部肌肉、輕盈的骨骼結構和特化的腳部。奔跑時,這些鳥類通常採用類似於哺乳動物的跑步姿態,身體前傾,翅膀微微展開以保持平衡。
跳躍是一些小型地棲鳥類常用的運動方式,特別是在複雜的地形中。例如,許多雀形目(Passeriformes)的地棲物種如畫眉(Laughingthrush)和鵪鶉(Quail)常常通過一系列快速的小跳來移動。這種運動方式要求鳥類具有強健的腿部肌肉和靈活的關節,能夠快速蹬地和著陸。跳躍不僅用於日常移動,也是一些物種躲避捕食者的重要策略。
攀爬雖然不是所有地棲鳥類都具備的能力,但在一些特定的生態位置中扮演著重要角色。例如,攀雀(Woodcreeper)雖然主要在地面活動,但能夠靈活地在樹幹上攀爬尋找食物。這種運動方式要求鳥類具有特化的腳爪結構和強壯的腿部肌肉,能夠牢固地抓住樹皮或岩石表面。
地棲鳥類的陸地運動方式與其特定的形態特徵密切相關。首先是腿部結構的適應。與飛行鳥類相比,地棲鳥類通常有更長、更強壯的腿,這不僅提供了更強的支撐力,也增加了步幅。腳部結構也有特殊適應,例如鴕鳥只有兩個腳趾,這減少了與地面的摩擦,有利於高速奔跑。
骨骼結構的改變也是地棲鳥類適應陸地生活的重要方面。許多物種的胸骨龍骨(Keel)退化,反映了飛行肌肉的減少和陸地運動肌肉的增強。同時,為了支撐更大的體重,地棲鳥類的骨骼通常更加堅固,尤其是支撐腿部的骨骼。
肌肉系統的適應同樣顯著。地棲鳥類的腿部肌肉高度發達,不僅提供了強大的推進力,還能夠支持長時間的運動。例如,鴕鳥的腿部肌肉佔其總體重的三分之一左右,這使得它們能夠保持長時間的高速奔跑。
平衡能力是地棲鳥類陸地運動的另一個關鍵要素。雖然失去了飛行能力,但許多物種仍保留了翅膀,並將其用於平衡。例如,鴕鳥在高速轉彎時會展開翅膀輔助平衡。此外,地棲鳥類通常具有發達的前庭系統,能夠精確感知身體的位置和姿態變化,這對於在不平坦的地形上移動至關重要。
地棲鳥類的陸地運動還涉及到複雜的能量平衡問題。陸地運動,特別是高速奔跑,需要消耗大量能量。因此,這些鳥類通常具有高效的心血管系統和呼吸系統。例如,鴕鳥的心臟相對體型來說特別大,能夠支持高強度的運動。同時,許多地棲鳥類還發展出了特殊的散熱機制,如鴕鳥的翅膀可以幫助調節體溫。
感知能力對地棲鳥類的陸地運動也極為重要。這些鳥類通常具有優秀的視力,能夠在奔跑中保持對周圍環境的清晰感知。一些物種如鴕鳥的眼睛位置較高,提供了更好的視野。聽覺同樣重要,許多地棲鳥類能夠通過聲音辨識潛在的威脅或獵物。
地棲鳥類的陸地運動還體現在它們的社會行為中。許多物種如火雞和雉雞會形成集體活動的群體。在這些群體中,個體需要協調自己的運動以保持群體的凝聚力。這種群體運動不僅有助於防禦捕食者,還可能提高覓食效率。
棲息地的特性對地棲鳥類的陸地運動有顯著影響。開闊草原上的物種如鴕鳥和班鳩(Sandgrouse)通常發展出了高速奔跑的能力。而生活在森林或灌木叢中的物種如松雞(Grouse)則更依賴於敏捷的跳躍和快速的方向變換。一些生活在岩石地帶的物種如岩雷鳥(Ptarmigan)則發展出了在陡峭地形上穩定移動的能力。
研究地棲鳥類的陸地運動對於理解陸地生態系統的功能具有重要意義。這些鳥類往往是生態系統中重要的種子傳播者和昆蟲控制者。通過它們的運動模式,我們可以更好地理解生態系統中能量和物質的流動。
此外,地棲鳥類的陸地運動研究對生物力學和機器人技術也有重要啟示。例如,鴕鳥的高效奔跑機制已經inspire了一些雙足機器人的設計。研究這些鳥類如何在複雜地形中保持平衡和靈活運動,可能為開發更先進的陸地機器人提供借鑒。
隨著技術的進步,研究地棲鳥類陸地運動的方法也在不斷創新。高速攝影和三維運動捕捉技術使得研究者能夠詳細分析這些鳥類的運動力學。GPS追蹤技術則提供了關於這些鳥類在自然環境中長期運動模式的寶貴數據。此外,計算機模擬技術也被廣泛應用於研究地棲鳥類的運動機制和能量消耗。
地棲鳥類的陸地運動能力是長期進化的結果,反映了生物對陸地環境的精細適應。這些鳥類通過形態、生理和行為上的一系列適應,成功地在地面生態位置中建立了自己的生存策略。研究它們的陸地運動不僅能夠幫助我們更好地理解生物適應性的奧秘,也為解決人類在陸地運動和機器人技術方面面臨的挑戰提供了新的思路。通過深入研究地棲鳥類的陸地運動,我們可以更全面地認識陸地生態系統的複雜性,為生物多樣性保護和可持續發展提供科學依據。
5.20樹棲鳥類的陸地運動
樹棲鳥類是一群適應在樹上生活的鳥類,牠們的身體結構和行為模式都經過長期演化以適應樹上生活。然而,即使是高度適應樹棲生活的鳥類,有時也需要在地面上活動。這種陸地運動對於樹棲鳥類來說是一種挑戰,因為牠們的身體結構主要是為了在樹枝間靈活移動而設計的。
樹棲鳥類在地面上的運動方式與其他地棲鳥類有所不同。大多數樹棲鳥類在地面上的移動顯得笨拙且緩慢,這主要是因為牠們的腳部結構和姿勢不適合在平坦的地面上行走。樹棲鳥類通常具有較長的腳趾和彎曲的爪子,這些特徵有助於牠們緊緊抓住樹枝,但在地面上卻不太實用。
許多樹棲鳥類,如鸚鵡科(Psittacidae)的成員,在地面上移動時會採用一種獨特的搖擺步態。這種步態涉及將身體從一側搖擺到另一側,同時向前移動。這種運動方式雖然看起來不太優雅,但實際上是一種有效的適應,能夠彌補牠們在地面上行走時的不足。
一些樹棲鳥類,如啄木鳥科(Picidae)的成員,已經發展出一種特殊的運動方式,稱為「跳躍式行走」。這種方式涉及使用雙腳同時跳躍,而不是像許多地棲鳥類那樣交替使用單腳行走。這種跳躍式的移動方式使啄木鳥能夠在地面上更快速、更有效地移動,儘管牠們的身體結構主要適應於在樹幹上攀爬。
值得注意的是,一些樹棲鳥類,如某些鴿子和鳩子科(Columbidae)的成員,已經進化出在地面和樹上都能靈活移動的能力。這些鳥類在地面上的運動能力相對較強,能夠有效地在地面上覓食和活動,同時仍保持在樹上棲息和築巢的能力。
樹棲鳥類在地面上的活動通常與幾個重要的生存需求有關。首先是覓食行為,許多樹棲鳥類會下到地面尋找食物,特別是當樹上的食物來源不足時。地面上可能有豐富的種子、昆蟲或其他食物資源,這些都是樹棲鳥類的重要營養來源。
其次,飲水也是樹棲鳥類需要下到地面的一個重要原因。雖然有些鳥類可以從樹葉或樹洞中收集到水分,但大多數鳥類還是需要定期到地面的水源處飲水。這種行為不僅滿足了生理需求,也為鳥類提供了社交互動的機會,因為飲水點往往是不同鳥類聚集的地方。
此外,一些樹棲鳥類會在地面上進行沙浴或泥浴。這種行為有助於維護羽毛的健康,可以去除寄生蟲,調節羽毛中的油脂含量,並保持羽毛的保暖和防水功能。雖然這種行為使樹棲鳥類暴露在地面捕食者的威脅之下,但其重要性使得許多鳥類仍然冒險這麼做。
在繁殖季節,一些樹棲鳥類可能需要在地面上收集築巢材料。雖然大多數材料可以直接從樹上獲得,但某些特殊的材料,如泥土或特定的植物纖維,可能需要鳥類下到地面才能獲取。這種行為再次說明瞭樹棲鳥類如何在適應樹上生活的同時,也保留了在必要時利用地面資源的能力。
然而,在地面上活動對樹棲鳥類來說並非沒有風險。地面是許多捕食者的活動區域,如貓科動物、蛇類和某些哺乳動物。樹棲鳥類在地面上的移動速度較慢,反應可能不如在樹上靈敏,這使得牠們更容易成為捕食者的目標。因此,許多樹棲鳥類在地面活動時會表現得特別警惕,經常抬頭觀察周圍環境,並在感到威脅時迅速飛回樹上。
為了應對地面活動的風險,一些樹棲鳥類已經發展出特殊的行為策略。例如,群居的樹棲鳥類可能會採用「哨兵」系統,即在群體中有一兩隻鳥停留在較高的樹枝上警戒,而其他成員在地面覓食。這種合作行為大大提高了群體在地面活動時的安全性。
某些樹棲鳥類,如某些鸚鵡物種,甚至能夠使用喙和腳配合,在地面上進行複雜的操作。例如,牠們可以用喙和腳配合來剝開堅果或處理其他食物。這種能力說明瞭即使是高度適應樹棲生活的鳥類,也能夠在地面上展現出令人驚訝的靈活性和適應性。
研究樹棲鳥類的陸地運動對於理解鳥類的行為生態學和進化生物學具有重要意義。它揭示了物種如何在不同環境之間平衡適應,以及如何在保持主要生態位的同時,仍能利用其他生態位的資源。這種適應性不僅體現在身體結構上,還體現在行為策略的靈活性上。
隨著城市化的推進,許多樹棲鳥類面臨著新的挑戰。都市環境中的人工結構,如建築物和人行道,為這些鳥類提供了新的「地面」環境。一些樹棲鳥類已經顯示出適應這種新環境的能力,在城市中的人工結構上覓食和活動。這種適應性再次強調了樹棲鳥類在面對環境變化時的靈活性。
樹棲鳥類的陸地運動這一主題還引發了關於鳥類運動能力進化的更廣泛討論。它提出了一個有趣的問題:是否所有的鳥類都起源於樹棲祖先,然後某些類群逐漸適應了地面生活,還是地棲能力是鳥類的原始特徵,而樹棲能力是後來進化而來的這個問題的答案可能涉及對早期鳥類化石記錄的深入研究,以及對現代鳥類運動能力的詳細比較分析。
5.21食肉猛禽的陸地運動
食肉猛禽是一群適應捕食生活的鳥類,包括鷹、隼、鴞等。這些鳥類以其強大的飛行能力、銳利的視力和有力的爪子而聞名,主要依靠飛行來捕獵和移動。然而,儘管牠們在空中表現出色,食肉猛禽在某些情況下也需要在地面上活動。這種陸地運動雖然不是牠們的主要活動方式,但在牠們的生存和行為中扮演著重要角色。
食肉猛禽的陸地運動能力因種類而異,但總體來說,相較於專門的地棲鳥類,牠們在地面上的活動顯得不那麼靈活。這主要是因為牠們的身體結構主要適應於飛行和抓取獵物。食肉猛禽通常具有強壯的腿部和鋒利的爪子,這些特徵在飛行和捕獵時非常有用,但在地面移動時可能成為限制因素。
大多數食肉猛禽在地面上採用一種獨特的步行方式,通常被描述為「蹣跚」或「笨拙」的步態。這種步態涉及將身體重心從一側移到另一側,同時向前移動。雖然這種移動方式看起來不太優雅,但實際上是一種有效的適應,能夠在保持平衡的同時,使猛禽能夠在地面上進行必要的活動。
一些較大型的猛禽,如某些鷹科(Accipitridae)的成員,在地面上的移動能力相對較好。例如,禿鷹(Vultures)經常需要在地面上行走以接近和處理大型動物屍體。這些鳥類已經進化出相對較強的腿部肌肉和更適合行走的腳掌結構,使牠們能夠在地面上更有效地移動。
相比之下,一些專門的空中獵手,如隼科(Falconidae)的成員,在地面上的活動能力較弱。這些鳥類通常避免長時間在地面上停留,只有在必要時才會短暫地降落在地面上。
食肉猛禽在地面上的活動通常與幾個關鍵的生存需求有關。首先是捕食行為。雖然大多數猛禽偏好從空中俯衝捕獵,但有時也需要在地面上追捕或處理獵物。例如,當獵物躲藏在灌木叢中或地面的凹陷處時,猛禽可能需要降落並在地面上進行最後的捕獲。在這種情況下,猛禽的地面運動能力就顯得尤為重要。
其次,食肉猛禽有時需要在地面上處理較大型的獵物。例如,當一隻鷹捕獲了一隻兔子或其他中型哺乳動物時,它可能需要在地面上停留一段時間來撕裂和食用獵物。在這個過程中,猛禽需要保持平衡,同時使用喙和爪子來處理食物。這種行為要求猛禽具備一定的地面運動能力和穩定性。
此外,飲水也是食肉猛禽需要在地面活動的一個重要原因。雖然許多猛禽能夠從獵物中獲得大部分所需的水分,但在乾燥的環境中,牠們可能需要定期前往水源飲水。這種行為不僅滿足了生理需求,也為猛禽提供了清潔羽毛的機會。
在繁殖季節,一些食肉猛禽可能需要在地面上收集築巢材料。雖然大多數材料可以直接從空中獲取或在樹上收集,但某些特殊的材料可能需要鳥類降落到地面才能獲得。這種行為再次強調了食肉猛禽在適應空中生活的同時,也保留了在必要時利用地面資源的能力。
值得注意的是,某些食肉猛禽,特別是那些適應了特定生態位的種類,可能比其他種類更依賴地面活動。例如,草原上的貓頭鷹,相比於森林中的近親,可能更經常在地面上活動。這些鳥類已經發展出更適合在開闊地形中行走和跑動的能力,反映了牠們對特定棲息地的適應。
然而,在地面上活動對食肉猛禽來說並非沒有風險。地面是許多其他捕食者的活動區域,如狐狸、郊狼等哺乳動物。食肉猛禽在地面上的移動速度較慢,反應可能不如在空中靈敏,這使得牠們更容易成為其他掠食者的目標。因此,許多食肉猛禽在地面活動時會表現得特別警惕,經常抬頭觀察周圍環境,並在感到威脅時迅速飛離。
為了應對地面活動的風險,一些食肉猛禽已經發展出特殊的行為策略。例如,某些群居的禿鷹在進食時會採用「輪班」系統,即在群體中有一些個體保持警戒,而其他成員專注於進食。這種合作行為大大提高了群體在地面活動時的安全性。
食肉猛禽的陸地運動能力在不同的年齡階段也有所不同。幼鳥在學習飛行之前,可能需要更多地依賴地面運動。這個階段的猛禽通常表現出較強的地面活動能力,能夠在巢區附近的樹枝或地面上移動。隨著飛行能力的發展,牠們對地面運動的依賴逐漸減少。
研究食肉猛禽的陸地運動對於理解這些鳥類的行為生態學和進化生物學具有重要意義。它揭示了物種如何在不同環境之間平衡適應,以及如何在保持主要生存策略的同時,仍能利用其他環境資源。這種適應性不僅體現在身體結構上,還體現在行為策略的靈活性上。
隨著人類活動的擴張,許多食肉猛禽面臨著新的挑戰和機遇。都市環境為這些鳥類提供了新的「地面」環境,如建築物屋頂和停車場。一些適應性較強的猛禽種類,如某些鷹和隼,已經顯示出適應這種新環境的能力,在城市中的人工結構上覓食和活動。這種適應性再次強調了食肉猛禽在面對環境變化時的靈活性。
食肉猛禽的陸地運動能力也引發了關於鳥類運動能力進化的更廣泛討論。它提出了一個有趣的問題:現代猛禽的地面運動能力是原始特徵的殘留,還是為了適應特定生態位而重新進化的結果?這個問題的答案可能涉及對早期猛禽化石記錄的深入研究,以及對現代猛禽運動能力的詳細比較分析。
食肉猛禽的陸地運動雖然不如牠們的飛行能力那樣引人注目,但在牠們的生存策略中扮演著不可或缺的角色。這種能力反映了自然界中生物適應性的複雜性和多樣性,同時也為我們理解鳥類運動能力的演化提供了寶貴的見解。
5.22貓頭鷹的夜間運動
貓頭鷹是夜行性猛禽中的佼佼者,牠們的夜間運動能力堪稱鳥類世界中的奇蹟。這些神秘的生物經過長期演化,發展出一系列獨特的適應性特徵,使牠們能在黑暗中自如地活動和捕獵。貓頭鷹的夜間運動涉及多個方面,包括飛行、獵捕、覓食以及在樹枝或地面上的活動,每個方面都體現了貓頭鷹對夜間生活的精妙適應。
貓頭鷹的夜間飛行能力首先得益於牠們特殊的羽毛結構。貓頭鷹的飛羽邊緣具有鋸齒狀結構,這種設計能夠顯著減少空氣流動產生的噪音。此外,貓頭鷹的羽毛表面覆蓋著一層細小的絨毛,進一步降低了飛行時的聲音。這些特徵使貓頭鷹能夠悄無聲息地在夜空中滑翔,不驚動潛在的獵物,也不引起其他掠食者的注意。
貓頭鷹的眼睛是牠們夜間視力的關鍵。相對於體型而言,貓頭鷹擁有極大的眼睛,這使得牠們能夠在微弱的光線下收集更多的光線。貓頭鷹的視網膜上分佈著大量的感光細胞,特別是感光棒細胞,這些細胞在弱光條件下特別有效。此外,貓頭鷹的眼睛還具有一種反光層,稱為眼底泡(Tapetumlucidum),能夠反射進入眼睛的光線,進一步增強視覺效果。這些特徵使貓頭鷹能夠在人類看來幾乎完全黑暗的環境中辨識物體。
然而,貓頭鷹的夜間運動並不僅僅依賴於視覺。牠們的聽覺系統同樣發達,在某些情況下甚至比視覺更為重要。貓頭鷹的耳朵位於頭部兩側,而且左右耳朵的位置略有不同。這種非對稱的結構使貓頭鷹能夠精確地定位聲音的來源。當獵物在地面上移動時,即使是極其微弱的沙沙聲,貓頭鷹也能準確地判斷其位置。
貓頭鷹特有的面部盤羽結構也在聲音定位中發揮重要作用。這些羽毛形成了一個類似衛星盤的結構,能夠收集和引導聲波到耳朵。這種設計進一步增強了貓頭鷹的聽覺能力,使牠們能夠在完全黑暗的環境中僅憑聲音就能成功捕獲獵物。
在夜間獵捕過程中,貓頭鷹展現出令人驚嘆的飛行技巧。牠們能夠在空中快速轉向,甚至能夠在狹窄的空間中靈活操縱。這種飛行能力部分歸功於貓頭鷹的翅膀結構。貓頭鷹的翅膀相對較短且寬闊,這種設計使牠們能夠在低速飛行時保持良好的控制力,同時也有助於在密集的樹林中穿梭。
貓頭鷹的獵捕行為通常涉及幾個階段。首先,貓頭鷹會利用其敏銳的視聽覺探測潛在的獵物。一旦發現目標,貓頭鷹會保持靜止,仔細觀察和聆聽以確定獵物的確切位置。然後,貓頭鷹會悄無聲息地接近獵物,通常是通過滑翔飛行。在最後的攻擊階段,貓頭鷹會突然加速,伸出強壯的爪子抓住獵物。
貓頭鷹的爪子是牠們夜間捕獵的另一個關鍵工具。這些爪子不僅鋒利,而且力量驚人。貓頭鷹的腳趾能夠靈活地調整抓握力度,既能輕柔地抓住小型獵物,也能有力地控制較大的獵物。這種精準的控制力使貓頭鷹能夠在各種不同的環境中成功捕獵,從開闊的草原到茂密的森林。
除了飛行和捕獵,貓頭鷹的夜間運動還包括在樹枝或地面上的活動。雖然貓頭鷹主要是飛行性鳥類,但牠們也需要在固定物體上停留,無論是為了休息、觀察還是處理獵物。貓頭鷹的腳部結構適應了這種需求,能夠牢固地抓住樹枝或其他棲息地。一些貓頭鷹種類,如倉鴞(BarnOwl),甚至能夠在地面上靈活移動,這在追捕地面獵物時特別有用。
貓頭鷹的夜間覓食行為也值得關注。不同種類的貓頭鷹有不同的食性和獵捕策略。例如,某些小型貓頭鷹主要捕食昆蟲,而大型種類則可能獵捕小型哺乳動物甚至其他鳥類。這種多樣性反映了貓頭鷹如何適應不同的生態位,並在夜間食物鏈中扮演重要角色。
貓頭鷹的夜間活動還涉及社交和交流。儘管大多數貓頭鷹種類都是獨居動物,但在繁殖季節,牠們會發出各種聲音來吸引配偶或宣示領地。這些聲音通常在夜間傳播得更遠,是貓頭鷹夜間行為的重要組成部分。
值得注意的是,並非所有的貓頭鷹都完全是夜行性的。一些種類,如短耳鴞(Short-earedOwl),也會在黃昏或清晨活動。這種行為模式的差異反映了不同貓頭鷹種類對特定生態環境的適應。
貓頭鷹的夜間運動能力也使牠們在生態系統中扮演著獨特的角色。作為夜間捕食者,貓頭鷹有助於控制小型哺乳動物和其他夜行性生物的數量,從而維持生態平衡。牠們的存在也為研究夜間生態系統提供了寶貴的機會。
然而,人類活動對貓頭鷹的夜間行為造成了一定影響。光污染是一個主要問題,過度的人工照明可能幹擾貓頭鷹的自然行為模式,影響牠們的獵捕效率和繁殖成功率。此外,棲息地破壞和環境變化也對貓頭鷹的生存構成威脅。
研究貓頭鷹的夜間運動不僅有助於我們更好地理解這些神奇的生物,也為生物力學、聲學和視覺科學等領域提供了寶貴的見解。例如,貓頭鷹的靜音飛行技術已經啟發了飛機設計師開發更安靜的飛行器。貓頭鷹的聽覺系統也為開發更高效的聲音定位技術提供了靈感。
貓頭鷹的夜間運動能力是自然界中適應性進化的典範。這些能力不僅使貓頭鷹成為高效的夜間獵手,也使牠們在各種生態系統中扮演著不可或缺的角色。通過研究貓頭鷹的夜間行為,我們不僅能夠更深入地瞭解這些神秘的生物,還能獲得對整個生態系統運作的新見解。