


第二章 生物圈的演變
生命是地球的特徵。生物圈與地球的直徑有關,是位於對流層上部和多孔岩石和沉積物表層之間的一層極薄的生命維持層,在太空中可以清楚地看到;它是地球衛星照片上的藍色和綠色的原因。
所有已知的生命形式都建立在核酸-蛋白質系統的基礎上,儘管涉及不同化學成分的生命系統理論上是可能的。地球上的生命似乎在條件允許的情況下迅速發展起來。
高溫原始地球的冷卻是一個重要因素。在一個以極端溫度為標準的宇宙中,大多數生命形式都被限制在0到100°C的相對狹窄的範圍內。
生物圈中的非生物元素被生命深深地塑造,就像生命被周圍的環境條件塑造一樣。生物圈隨著時間的推移而增長。七億年前,它是一個狹窄的、可能是不連續的帶,只包括海洋較淺的部分。如今,它已經深入大氣層和海洋深處,甚至侵入了多孔岩石中的微小空間。因此,從地球最深處的9.9公里到海洋深處,從地球表面的11.6公里到海洋深處,生命從地球的最深處延伸到了海洋。
即使在地球上最惡劣的環境中,凍幹的南極沙漠藻類也能在半透明岩石中的微小空間中找到避難所。這些岩石提供了避風的地方,聚集了太陽光,起到了溫室的作用,使生物活動每年可以進行幾個星期。在另一個極端,深海火山裂縫噴口中有嗜熱細菌(嗜熱細菌),在這些噴口中,海水在巨大的壓力下被加熱到極高的溫度。一些研究人員認為,這些噴口中存在的一些嗜熱微生物可以在300°C以上的溫度下存活。如果溫度遠低於沸點,它們就會死亡。
生命是通過進化過程改變的。進化是遺傳、遺傳變異和因個體數量超過可用資源而產生的競爭的必然結果。自然選擇的結果是允許某些性狀比其他性狀永存。幾十億年來,這個過程導致了生命形式的巨大多樣化。
生命史的特點是進化變化的加速和不可預測的滅絕時期,隨後往往是迅速的多樣化。對於其中一些趨勢和事件的原因,甚至是重要性,仍有許多爭論。也許目前最熱議的問題是關於滅絕和多樣化的理論。在20世紀70年代早期,進化生物學家史蒂芬·傑伊·古爾德和尼爾斯·埃爾德雷奇開發了一個稱為“間斷平衡”的模型,它描述並解釋了物種形成的某些方面。這一理論假設進化不是以穩定的速度發展,而是以爆發式的速度發展,因為短暫的快速進化變化伴隨著長期的相對進化停滯。
生物圈中有機和無機元素之間的相互依賴程度以及負反饋和正回饋機制在維持生命中的重要性日益得到承認。英國物理學家詹姆斯·洛夫洛克(James Lovelock)和美國微生物學家林恩·馬古利斯(Lynn Margulis)認為,由於生物圈的元素是如此相互依存和相互關聯的,生物圈可以被視為一個單獨的、自我調節的有機體,他們稱之為蓋亞。
蓋亞假說假設地球表面、海洋和大氣的物理條件已經變得適合生命,並且由生物群自己維持在這種狀態。證據包括地球表面相對��定的溫度,在過去的35億年裡,儘管來自太陽的能量增加了25%,地球表面的溫度仍然保持不變。過去5億年來地球海洋和大氣化學的驚人的��定性也被用來支持這一理論。
蓋亞假說的一個重要組成部分是生物群參與生命中各種重要元素的循環。生物在碳和硫循環中所起的作用是生物活動的重要性以及生物圈中有機和無機元素之間複雜相互關係的一個很好的例子。
儘管蓋亞的概念提供了有趣的生物圈模型,但許多研究人員並不認為生物圈像蓋亞假說所暗示的那樣完全整合。
n 地球地質歷史與早期生命形式

據信,大氣的形成是由火山噴發釋放的氣體(放氣的一個例子)造成的。當地球表面冷卻時,新形成的大氣中的水蒸氣凝結成海洋的水。直到大約40億年前,海洋可能還太熱,無法維持生命。到28億年前,第一批輕質矽石和鋁質岩石已經形成,它們是典型的大陸。這些岩石迅速擴張,到26億年前,今天存在的大陸塊中有60%已經形成,允許大陸漂移的過程已經開始。
Lynn Margulis是一位美國生物學家,她的一系列關於真核細胞發育的內共生理論徹底改變了關於地球上生命起源的現代概念。
在她職業生涯的大部分時間裡,瑪古麗絲被那些追求傳統達爾文“適者生存”的生物學方法的同齡人視為激進者。她的觀點側重于共生,即兩種不同的有機體在一種既有利也不利的組合中的生活安排,經常遭到懷疑甚至敵意的攻擊。她最重要的工作之一是細胞起源的系列內共生理論(SET)的發展,該理論認為真核細胞(有核細胞)是由先前獨立存在的無核細菌的共生合併演化而來的。在這個理論中,線粒體和葉綠體是真核細胞的兩個主要細胞器,是曾經自由生活的細菌的後代。她在她的第一本書《真核細胞的起源》(1970)中解釋了這個概念。當時,她的理論被認為牽強附會,但後來被廣泛接受。她在1981年的經典著作《細胞進化中的共生》中詳細闡述了這一次細胞與細菌的另一種共生合併,螺旋體是一種波動迅速發展成有核細胞內部運輸系統的細菌。Margulis進一步推測真核纖毛也是螺旋體,細胞質是由真細菌和古細菌的共生關係演化而來。
她1982年與美國生物學家卡琳(Karlene V.Schwartz)合著的《五領域》(Five Kingdoms)一書闡述了地球生命分類的五領域體系—動物、植物、細菌(原核生物)、真菌和原生動物。原生生物界包含了其他系統中大多數單細胞生物(和多細胞藻類),但由於過於籠統而遭到拒絕。許多通常被歸類為原生生物的有機體被置於另外四個領域中的一個;原生動物構成了其餘的有機體,它們都是水生的,包括藻類和黏菌。馬古利斯編輯了原生動物手冊(1990年)綱要的部分內容。
另一個令瑪古利斯感興趣的領域是她與英國科學家詹姆斯洛夫洛克(jameslovelock)在有爭議的蓋亞假說上的長期合作。這一觀點認為,地球可以被視為一個自我調節的有機體,也就是說,一個複雜的實體,其生命體和無機元素相互依存,其生命形式積極地改變環境,以維持適宜的條件。
生命的發展
最古老的無可爭議的化石大約有35億年的歷史。生命似乎起源於39至35億年前。形成生命所需的基本化學成分在地球上和已知宇宙的其他地方都很豐富。生命最初可能是通過有機小分子的自我組裝而產生的。在這個過程中,粘土或晶體的表面可能起到了範本的作用。脫水和冷凍也可能在更複雜分子的組裝過程中發揮了作用。
在20世紀50年代,斯坦利·公尺勒和哈樂德·烏雷在芝加哥大學進行了一系列著名的實驗,在太古宙Eon(3.8億到25億年前)。電火花代替閃電,被引入到氣體混合物中,這些氣體反應形成氨基酸,氨基酸是蛋白質的基本組成部分。後來的實驗產生了構成DNA結構的核苷酸堿基。這些基本的積木是如何組合起來形成生命的仍不清楚。這個過程可能花費了數百萬年。
生命的起源
化石記錄中最早的簡單生命形式是原核生物(沒有被膜包圍的細胞核的細胞有機體),即細菌和藍藻(以前稱為藍綠藻)。它們在被稱為疊層石的岩石中被發現,疊層石是層狀的,球狀的,通常是鈣質的,通常比足球還大。疊層石是原核生物群體被困在沉積物中形成的;它們是很容易辨認的化石,對該領域的研究人員來說是顯而易見的。對疊層石化石的薄層切片偶爾會發現製造疊層石的微生物的顯微化石細胞。
直到大約25到28億年前,地球的大氣主要由二氧化碳組成。由於原始細菌和藍藻通過光合作用或相關的生命過程捕獲大氣中的碳,並將其沉積在海底,因此碳從大氣中被清除。通過可能與板塊構造有關的地質作用,這些碳被帶入地殼。目前,每年約有0.1%的固定碳以這種方式流失到生物圈中。在元古宙Proterozoic(25億至5.42億年前),這一過程使一些自由氧首次存在於大氣中。
藍綠藻(菌)也是第一個在光合過程中利用水作為電子和氫的來源的生物。這種反應釋放出游離氧,並開始在大氣中積累,使依賴氧的生命形式得以進化。
日益增長的複雜性
1992年在美國密歇根州發現的化石表明,第一批真核生物出現在大約20億年前。這些複雜的單細胞生物,如變形蟲,與原核生物不同的是,它們有一個膜結合的細胞核,成對的染色體,在大多數情況下,還有線粒體。它們也需要氧氣才能發揮作用。
生物圈演化的重大變化發生在前寒武紀晚期(約7億至5.42億年前)。在此之前,單細胞真核生物首次出現後的14億年裡,它一直是地球上的主要生命形式。然後,在前寒武紀晚期,複雜的多細胞生物(由大量或多或少特殊化的細胞組成的動物或植物)迅速進化和多樣化。
生命發展面臨的挑戰
在此之前,複雜生命的發展可能受到生物群所產生的大氣變化的阻礙。以前大氣中大量的二氧化碳產生了一種絕緣或溫室效應。當有機體將這種氣體從大氣中排出時,溫室效應就減輕了,地球的氣候也發生了變化。據信,這一事件導致了嚴重的冰河期席捲地球。
冰河時代的成因仍在激烈爭論中。地質學家約翰·詹姆斯·韋弗斯(John james veevers)在1990年提出了一個假說,認為它們的出現與大陸漂移有關。根據這一模型,大陸漂移是週期性的:在過去的12億年裡,大陸在地球上所有大陸塊都是分開的階段和這些不同的大陸形成一個大陸的“超級大陸”階段之間波動。在超大陸階段,海底幾乎沒有擴張,同時伴隨著從地幔釋放二氧化碳。這樣,大氣中的二氧化碳就會減少,溫室效應也會減輕,從而創造一個更涼爽的環境。據信,主要冰期與每一個超大陸相一致。(然而,過去兩百萬年的冰河期是短暫的、振盪的,被認為不是這個更大週期的一部分。)
依賴於附近海岸線或陸地棲息地的生命形式的分佈受到大陸相對位置的影響。超大陸的週期性分裂為進化提供了許多孤立的機會。今天的澳大利亞是最孤立的大陸,其獨特的動植物群是眾所周知的。在過去,其他大陸即使不是更加孤立,也同樣如此。現在中亞的一部分,被稱為哈薩克斯坦,是一塊孤立的大陸,存在於寒武紀後半期(大約5.13億年前)和泥盆紀中期(約3.85億年前)之間。在這些大陸和其他大陸上,形成了獨特的植物群和動物群。
溫室效應
溫室效應是用來描述地球表面和對流層(大氣最底層)變暖的一種表達方式,它是由空氣中的水蒸氣、二氧化碳、甲烷和某些其他氣體引起的。在這些被稱為溫室氣體的氣體中,水蒸氣的影響最大。
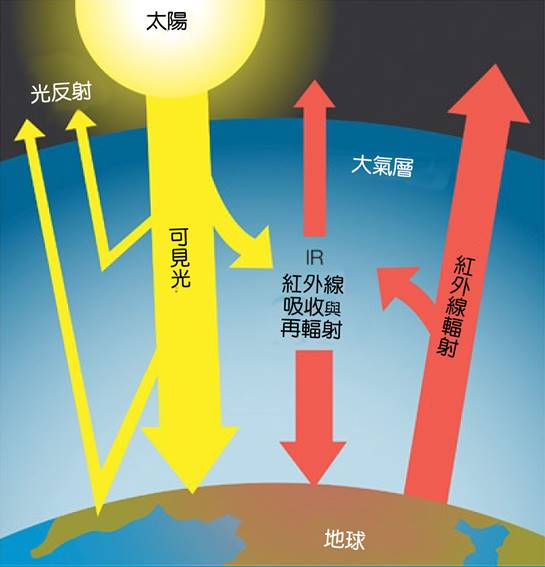
地球上的溫室效應。一些入射的太陽光被地球的大氣層和地表反射,但大部分被地球表面吸收,而地表被加熱了。然後從表面發射紅外輻射。一些紅外輻射會逃逸到太空,但有些會被大氣中的溫室氣體(特別是水蒸氣、二氧化碳和甲烷)吸收,然後向各個方向再輻射,有些輻射到太空,另一些則返回地表,在那裡它進一步加熱了地表和低層大氣. 大英百科全書公司。
大氣允許來自太陽的大部分可見光通過並到達地球表面。當地球表面被太陽光加熱時,它將部分能量以紅外線輻射的形式輻射回太空。與可見光不同,這種輻射往往被大氣中的溫室氣體吸收,從而提高其溫度。加熱的大氣反過來又向地球表面輻射紅外輻射。(儘管它的名字叫“溫室效應”,但它不同於溫室裡的溫室效應,溫室裡的玻璃窗傳遞可見的陽光,但通過捕捉溫暖的空氣來保持室內的熱量。)
如果沒有溫室效應引起的加熱,地球的平均表面溫度只有-18°C(0°F)。在金星,大氣中高濃度的二氧化碳會引起極端的溫室效應,導致地表溫度高達450°C(840°F)。
雖然溫室效應是一種自然發生的現象,但由於人類活動,溫室氣體排放到大氣中,溫室效應有可能加劇。從工業革命開始到20世紀末,大氣中二氧化碳的含量增加了30%,甲烷的含量增加了一倍多。一些科學家預測,與人類有關的大氣二氧化碳和其他溫室氣體的增加可能導致全球平均氣溫在21世紀末上升1.4至5.8°C(2.5至10.4°F)。全球變暖可能改變地球的氣候,從而產生新的模式和極端的乾旱和降雨,並可能擾亂某些地區的糧食生產。然而,參與氣候研究的其他科學家認為,這種預測被誇大了。
滅絕與多樣化
直到20世紀80年代,早期多細胞生物的化石記錄被解釋為簡單而迅速的多樣化。古生物學家Adolf Seilacher和其他人認為這是不正確的,最早的多細胞生物群包括很少或根本沒有與後來的動物群直接祖先的物種。作為證據,他們指出了埃迪卡拉紀時期早期的動物群—生活在前寒武紀末期,距今6.35億年至5.42億年前,它們以南澳大利亞的埃迪卡拉山命名。據信,埃迪卡拉紀Ediacaran動物群很少與加拿大西部伯吉斯葉岩的晚期動物群有關(寒武紀中期[約5.2億至5億年前])。從這一觀點來看,化石記錄被認為是多細胞生物至少兩次或多或少獨立的進化輻射導致的嚴重滅絕。因此,即使在這個遙遠的生命滅絕時期,也會起到重要的作用。其他權威人士不同意這一模式,認為埃迪卡拉紀動物有顯生宙時代的親戚(包括古生代、中生代和新生代[5.42億年前至今]),例如海棚和多毛類蠕蟲。
寒武紀生命大爆發
寒武紀Cambrian的開始,現在被認為可以追溯到5.42億年前,而不是5.7億年前,見證了前所未有的生命爆炸。許多研究者認識到,在20到35歲之間的現代動物生活中,許多主要的門似乎是在那個時候進化而來的,可能只有幾百萬年的時間。在這段時間裡,許多其他的門進化了,其中絕大多數在接下來的5000萬到1億年間滅絕了。具有諷刺意味的是,許多最成功的現代門(包括脊索動物,包括所有脊椎動物)都是寒武紀集合中的稀有元素;包括節肢動物和海綿動物在內的門包含了寒武紀時期數量上最佔優勢的分類群(分類群),而這些都是滅絕的分類群。
寒武紀的開始以碳酸鈣殼等堅硬部分的演化為標誌。這些身體部位比軟組織更容易成為化石,因此化石記錄在它們出現後變得更加完整。許多動物譜系幾乎同時獨立進化出堅硬的部分。其原因仍在爭論中,但一個主要的理論是,大氣中的氧氣含量最終達到了允許大型複雜動物生存的水準。氧水準也可能促進了代謝過程中產生膠原蛋白,膠原蛋白是構成人體硬結構的基礎。
早寒武紀(5.42億年至5.1億年前)發生的其他重大變化包括:在海底沉積物中挖洞的動物物種的發展,而不是躺在海底沉積物上的動物物種的發展,以及第一批碳酸鹽岩礁的演化,這些生物礁是由被稱為古脊椎動物的海綿狀動物建造的。
到寒武紀早期,生物圈仍然局限于世界海洋的邊緣;陸地上沒有發現任何生物(除了潮濕沉積物中可能存在的藍藻[以前稱為藍綠藻],中上層物種(生活在公海中的生物群)相對較少,海洋深處沒有生物居住。然而,海底淺層地區的生物已經非常多樣化。這種早期的水生生態系統包括相對較大的食肉動物畸形龍舌鰨、以沉積物為食的三葉蟲(早期節肢動物)和軟體動物、懸浮覓食海綿動物、各種食腐節肢動物,甚至可能包括甲蟲亞綱等寄生蟲。因此,在這個時候,一個發達的水生生態系統似乎已經在海洋淺水區運行。
寒武紀之後,生物圈繼續相對迅速地擴張。在奧陶紀時期(4.88億至4.44億年前),出現了典型的古生代海洋動物群,包括苔蘚蟲、腕足動物、珊瑚、鸚鵡螺和海百合。由於環境變化,許多海洋物種在奧陶紀末期滅絕。早志留紀(4.44億至4.23億年前)標誌著海洋中許多懸浮食性動物快速進化的時期。結果,像鸚鵡螺這樣的遠洋捕食者變得豐富起來。最古老的顱骨魚類,在晚志留世(4.23億至4.16億年前)變得常見。
陸地生命的出現
大約在4.2億年前,植物入侵了志留紀Silurian晚期的陸地,到了4.1億年前,陸地上發現了各種節肢動物。到了泥盆紀Devonian中期(大約3.9億年到3.8億年前),真正的蜘蛛已經進化出能夠吐絲的蜘蛛。大約5000萬年後,有翅膀的昆蟲緊隨其後。到了泥盆紀晚期(3.85億至3.59億年前),一些脊椎動物也出現在陸地上。它們產生了脊索動物、兩棲動物、爬行動物、鳥類和哺乳動物。
陸生植物被認為是由綠藻等綠藻植物進化而來的。它們在陸地上的生存需要特殊的適應,以防止它們乾涸,並幫助它們獲得營養和繁殖。角質在植物上形成蠟質層(角質層),氣孔有助於防止乾燥,根系和支援組織的發育有助於提供營養,孢子和種子提供了繁殖手段。
真菌是陸地植物的早期夥伴。菌根真菌似乎與諸如Rhynia(蕨類植物的可能祖先)、馬尾植物和種子植物的根有關,而類似苔蘚的植物碎片則保存在古代岩石中(地衣是真菌和藻類的共生組合)。
最早被保存下來的廣泛分佈的陸地植物,也是已知最古老的維管植物(一種具有特殊組織的植物,可以運輸水和營養物質並提供支援),是庫克索尼亞。這種祖先植物的結構類似苔蘚,在幾個大陸上有4.1億年歷史的岩石中被發現。庫克索尼亞,或類似的植物,很快就產生了維管植物的所有其他分支。一些最早的維管植物包括原石松屬和Rhynia,它們都是晚志留紀至早泥盆紀。到中泥盆世,一些植物譜系中形成層和韌皮部的發育使樹形物種得以發育。這種巨大的穴居動物是現代苔蘚類植物的近親,在這個時候特別豐富。種子或類似種子的結構很快就出現在許多植物譜系中。
為了適應陸地上的生活,最早的陸生脊椎動物面臨著與植物相似的問題。一組肉質鰭、呼吸空氣的魚類中的一些成員被認為是陸棲脊椎動物的祖先。尤瑟諾普頓是其中最著名的。
到泥盆紀Devonian晚期,最早的四足動物已經出現。像魚鱗蟲和刺五加(都來自格陵蘭東部)是最著名的。它們被恰當地描述為有腿的魚,被歸類為迷路齒兩棲動物,它們保留了許多類似魚的特徵,包括鰓、每英尺高達8個手指和尾鰭。
在泥盆紀時期,魚類發生了迅速的進化,所有的主要類群都在這一時期出現或多樣化。在這一時期最著名和最具特色的魚類是斑尾魚(已滅絕的有顎魚類)。許多是重甲物種,導致了底棲生物的存在,而其他物種是遠洋的,鱗片較輕。它們在泥盆紀末期滅絕了。
在石炭紀Carboniferous(3.59億至2.99億年前)和早二疊世Permian(2.99億至2.71億年前),迷路齒形動物成為主要的生命形式,進化成無數種。許多譜系在二疊紀末期(2.51億年前)滅絕,儘管至少有一種在南半球高緯度地區一直存在到白堊紀早期(1.46億至9960萬年前)。三疊紀Triassic早期(2.51億至2.46億年前),包括青蛙和蠑螈在內的利薩非人首次出現在化石記錄中。

n 進化的進程

地球上的生命具有滅絕和物種多樣化的特點。大陸重排、軌道變化、大氣化學變化、外星撞擊和其他因素都會導致環境變化。環境變化經常擾亂生態系統,並威脅到居住在生態系統中的生物的生存。如果環境變化緩慢,滅絕率相對較低。然而,在某些情況下,變化是迅速的,在相對較短的時間內,整個有機體的譜系都消失了。在這類事件的餘波中,進化加速,新物種崛起,承擔起角色,並佔據了那些已經過世的物種的棲息地。今天的世界,有著複雜的生命形式和無數的生態系統,是幾個時期滅絕和物種形成的結果。
二疊紀冰川與乾旱
石炭紀Carboniferous中期到早二疊世Permian之間的時間間隔是冰期延長的特徵。所有大陸被連接成一個超級大陸(盤古大陸Pangea),一個巨大的冰原覆蓋了現在的南極洲、澳大利亞南部、印度大部分地區、非洲南部和南美洲東部的大部分地區。在泥盆紀和早石炭紀(3.59億至3.18億年前)溫暖的沼澤中繁衍生息的巨型穴居動物因此消失了。取而代之的是現在已經滅絕的種子蕨類植物,即所謂的始祖鳥植物群變得豐富起來。在南部大陸,二疊紀以舌蕨屬植物為主。這些神秘的喬木和灌木可能產生了中生代(2.51億至6550萬年前)的主要植物群,甚至可能是開花植物。到了二疊紀末期,裸子植物(種子植物的種子缺乏覆蓋)如銀杏和早期針葉樹已經出現。到三疊紀早期,它們已經在其他植物無法忍受的乾燥環境中廣泛傳播。
二疊紀的結束標誌著也許是地球上有記載的最大的滅絕事件。總的來說,大約96%的海洋物種消失了,包括角珊瑚和板狀珊瑚、三葉蟲、廣翅目、多數鸚鵡螺類群、許多棘皮動物群以及許多腕足類和苔蘚蟲類。典型的滅絕程度是苔蘚蟲的命運。在許多早期的類群中,只有一個苔蘚蟲譜系(輪蟲)在二疊紀危機中倖存下來。苔蘚動物在中生代早期之前一直很罕見,在白堊紀時期(1.46億至6550萬年前)再次變得豐富,並一直持續到現代。與無脊椎動物相比,脊椎動物受此事件影響較小。
爬行動物的輻射
最早的爬行動物化石是在大約3.4億年前的石炭紀岩石中發現的。這些早期的爬行動物產生了synapsid爬行動物,在二疊紀變得豐富起來。突觸動物是陸地上的食肉動物,包括一些非常大的物種,如二甲蟲,這種動物有著細長的神經刺,沿著它們的背部形成一個“帆”。三疊紀晚期,有一類突觸動物,即類獸爬行動物,產生了哺乳動物。
原始的底足類爬行動物產生了兩個主要的類群,lepidosaurs(“鱗狀爬行動物”),包括蜥蜴和蛇,以及包括恐龍和鱷魚的爬行動物佔據主導優勢(“統治爬行動物”)。它們最早出現在石炭紀晚期,大約3億年前,之後的6000萬年裡,它們仍然很小,具有普遍的特徵。直到二疊紀大滅絕之後,它們才開始多樣化,並隨著其規模、豐度和多樣性的增加而主宰環境。
三疊紀時期(2.51億至2億年前)開始時相對溫暖潮濕,但隨著它的發展,條件變得越來越炎熱和乾燥。在這段時間裡,原始的lepidosaurs繁衍生息;蝶骨龍(其中唯一倖存的成員是紐西蘭的tuatara)特別豐富。蜥蜴出現在三疊紀,而蛇是在大約1.2億年前的白堊紀早期從巨蜥進化而來的。
從中三疊紀(2.45億年前到2.28億年前)一直到白堊紀末期,始祖龍一直主宰著陸地生命。最著名的古龍是恐龍,但翼龍(會飛的爬行動物)、鱷魚和鳥類也屬於這一類。
鳥類被認為是從原始的古龍目進化而來的。最早的鳥類化石證據是侏羅紀晚期烏鴉大小的始祖鳥。
裸子植物是通過暴露的種子或胚珠繁殖的任何維管植物,與種子被成熟的子房或果實包圍的被子植物或開花植物相反。裸子植物的種子是不可見的。然而,這些毬果和水果不一樣。在授粉過程中,未成熟的雄性配子或花粉粒,在毬果鱗片中篩選,直接落在含有未成熟雌配子的胚珠上,而不是像被子植物那樣落在花的成分(柱頭和心皮)上。此外,成熟時,毬果膨大,露出裸露的種子。
1825年,蘇格蘭植物學家羅伯特·布朗首次將裸子植物與被子植物區分開來。曾經,它們被認為是一個單一的種子植物綱,被稱為裸子植物,但分類學家現在傾向于識別現存裸子植物的四個不同的分類(和目):松樹目(松果目)、蘇鐵植物門(蘇鐵綱)、銀杏屬(銀杏屬),以及片麻岩(Gnetophyta)和裸子植物(Gnetophyta)—僅非正式地使用裸子植物這個術語。並不是所有的裸子植物都有著密切的聯繫,數億年來它們一直是不同的類群。目前共有82屬,947種。裸子植物分佈於世界各地,具有廣泛的緯度和縱向分佈範圍。

白堊紀生命的多樣性
白堊紀Cretaceous時期大型恐龍,如食肉恐龍、霸王龍、食草三角龍和蜥腳類阿拉莫龍是陸地上的主要形態。海洋生物包括一些無脊椎動物,如球狀有孔蟲和鈣質放射蟲,它們在侏羅紀時期(2億至1.46億年前)非常豐富。他們的遺骸第一次被鈣質軟泥覆蓋在海底,這對於對比不同地點的沉積岩年代非常有用。現代軟體動物群,如蛤蜊和食肉蝸牛,以及硬骨魚(大多數現代魚類的前身),都是最早出現的,而蛇頸龍、上龍和摩沙龍(goannas的最後一個巨型親屬)是主要的捕食者。翼龍是主要的大型飛行動物。裸子植物,如銀杏、蘇鐵和蕨類植物是優勢植物,儘管被子植物在這一時期末期變得越來越普遍。最早出現在侏羅紀的鳥類和在三疊紀進化的哺乳動物也存在,但它們是地球動物群的次要組成部分,與它們在新生代時期(6550萬年前至今)的優勢形成鮮明對比。似乎在這個時候,各種昆蟲群很可能迅速多樣化,以應對開花植物傳播所帶來的生態機遇。
氣候的影響
溫室條件的最大發展發生在白堊紀,可能與大氣中二氧化碳等溫室氣體的增加有關。在這段時間裡沒有極地冰蓋,北極圈和南極圈內的陸地都能夠支持多種多樣的動植物生活。海平面比現在高得多,大陸的低地部分形成了廣闊而淺的內海。這種棲息地支持各種大型雙殼類動物,如形成珊瑚礁的rudistid和1公尺長的貽貝狀伊諾瑟馬斯。
對新發現的北極和南極白堊紀動植物的研究揭示了有趣的差異和一些與溫室條件有關的異常現象。北極動物群以大型恐龍為主,它們被認為是遷徙的。小型吸熱動物,如哺乳動物,但小型放熱物種如蜥蜴不存在。南極動物群有著驚人的不同。小的,食草的,鳥臀恐龍,如亞特拉斯科龍是最豐富的動物群。海龜和肺魚也出現了,而最大的食肉動物是兩公尺高的異龍和晚期存活的迷路齒龍。恐龍和其他南極動物顯然沒有遷徙,他們必須忍受幾個月的接近冰凍的環境和完全的黑暗。他們是如何應對的仍不清楚。
白堊紀大滅絕
白堊紀在6550萬年前突然結束,發生了一次大規模的滅絕事件。恐龍、菊石和大多數貝倫目動物(都與魷魚和鸚鵡螺有關)、蛤蜊和齒形鳥類都滅絕了。事實上,在性成熟時達到成年體重約25公斤(55磅)的所有動物物種似乎都在這個時候消失了。小型生物如鈣質浮游生物、玻璃海綿、淡水魚和腕足動物的多樣性嚴重減少,月桂樹類群的裸子植物和被子植物也是如此。
這場世界大滅絕事件的起因仍在激烈辯論中。許多生物、氣候和地外因素已經被提出來解釋它。沃爾特和路易士·阿爾瓦雷斯(Luis Alvarez)大約在1980年提出的小行星理論假設,滅絕是地球與直徑約10至20公里(約6至12英里)的小行星相撞的結果。一般認為,撞擊導致大量顆粒物排放到高層大氣中,遮住了太陽,導致光合活性急劇降低,全球變冷。
小行星理論促進了人們對物種滅絕的普遍興趣。一些研究人員假設滅絕事件是週期性的,大約每2600萬年發生一次。儘管這一理論沒有被廣泛接受,但人們正在形成一種共識,即滅絕事件比以前認識到的更頻繁、更具災難性、更具變數。也越來越明顯的是,由於它們隨機影響各種物種的生存或滅絕,滅絕是決定進化方向的主要因素之一。
古近紀 Paleogene 早期的轉換
在白堊紀大滅絕中倖存下來的物種的進化和生態反應似乎很快。一種倖存的蕨類植物被認為在災難發生後的10000年內覆蓋了地球表面的90%。包括哺乳動物、鳥類、開花植物及其相關昆蟲、藤壺和苔蘚蟲在內的各種類群迅速多樣化。由於大陸遷移,動植物也發生了分化。
哺乳動物的崛起
在現代哺乳動物的三大類群中,產卵的單孔目動物和有袋類動物數量相對較少,在南部大陸最為成功。單孔目動物是現存哺乳動物中最原始的一種,鴨嘴獸和針鼴僅存兩種。第三類哺乳動物,胎盤哺乳動物,取得了最大的成功,產生了飛行形式(蝙蝠)、海洋物種(鯨魚、海妖和海豹),以及各種各樣的陸生形式。
胎盤哺乳動物的多樣化迅速。在古近紀時期恐龍滅絕後的幾百萬年(6550萬至2300萬年前),一些胎盤類群,如食草動物(產生有蹄類動物)的數量翻了兩番。胎盤的大小和生態範圍也增加了。在白堊紀大滅絕時,最大的胎盤並不比貓大。到古新世末期Paleocene Epoch(6550萬至5580萬年前),大約1000萬年後,體重超過800公斤的物種已經進化。到那時,哺乳動物已經多樣化,以填補主要的生態位,包括那些大型食草動物,食肉動物,食腐動物和更專業的類型。因此,在古新世,哺乳動物屬和科的進化最為迅速。在新生代的其餘時間裡,開花植物及其伴生昆蟲也迅速擴張。它們在不同大陸上隔離後,也分化成不同的植物群和動物群。

靈長類動物的出現
在白堊紀晚期出現的有胎盤哺乳動物中,靈長類動物是其中之一。大約5萬年前,一個小類人猿進化為一個小類人猿。200萬年前,人類從這一系進化而來。在20萬到10萬年前,現代人類(即智人Homo sapiens),已經進化;據信,他們第一次離開非洲-歐亞大陸的家園,大約4萬年前入侵澳大拉西亞。到11000年前,他們已經進入美洲,從而完成了對可居住大陸的殖民統治。
新生代氣候波動
晚白堊世大滅絕事件(Late Cretaceous extinction event),之後的一段時間,即新生代Cenozoic Era(6550萬年前至今),氣候波動具有普遍的降溫趨勢。始新世早期(5580萬至4860萬年前)是溫暖的;然而,到始新世末期(3390萬年前),世界溫度突然下降。在中新世開始時(2303萬年前),溫暖的條件又恢復了,只是在這個時代結束時(533萬年前)消失了。從那時起,氣候一直在振盪,在大約240萬年前冰河時代開始時達到頂峰,世界冰蓋多次進退。
在始新世後期(大約4000萬年前),南極洲開始形成大量的雪場和相關的冰川,這是自二疊紀以來首次出現在大陸上。冰蓋形成于1650萬至1300萬年前,其最迅速的發展發生在1480萬至1400萬年前。到600萬年前,一個巨大的冰蓋終於把東西南極洲連接起來了。
水生生物變化的影響
在後始新世,南大洋底層水溫急劇下降,下降了4至5℃(7∼9℉)。這似乎是由南極洲及其周圍海域日益隔絕的物理和熱造成的。在早中新世(2303萬至1597萬年前)之前,南極與南美洲之間的德雷克通道的開通和南極繞極流的建立,完成了這一隔離。這最終導致了南極底層水的發展,這種冷水、深、營養豐富,今天發源于南極洲,向北流到世界上所有主要海洋。南極底水以其獨特的營養承載能力,對海洋生物產生了深遠的影響。正是由於這種能力,人們認為它在最初建立時就導致了營養循環的重大變化。例如,它可能是磷蝦數量豐富的原因,也可能是大神秘鯨(濾食性鯨魚)進化的原因,這種鯨魚最早出現在漸新世(3390萬至2303萬年前)。
冰期
六百萬年前南極洲大面積冰蓋的形成導致海平面急劇下降。在它的高度,中新世冰期末期,南極洲的羅斯冰架延伸到其目前位置以北約300至400公里(約190至250英里)。這一事件孤立了地中海,在隨後的溫度波動中,地中海經歷了無數次的蒸發和再填充循環。由於這些變化,大約有100萬立方公里(約24萬立方英里)的鹽和石膏被從世界海洋中移走,現在埋在地中海下面的沉積沉積物中。這些事件使世界海洋的含鹽量比以前減少了約6%。這反過來又促進了全球氣候的冷卻,因為鹽度的降低提高了海洋的冰點。這促進了高緯度海冰的形成,也增強了地球表面的反射率(反照率)。
晚中新世(1161萬至533萬年前)和更新世的冰期似乎是由與早期冰期不同的事件造成的,例如石炭紀和早二疊世。
地球軸線的傾斜和進動或擺動以及地球軌道形狀的變化被認為是導致最近的冰期的原因,這些冰期大約每10萬年重複一次。
第四紀大滅絕
更新世期間,哺乳動物繼續多樣化,伴隨著局部的和較少的大範圍滅絕事件。然而,在更新世末期(5萬至1萬年前),滅絕事件發生時,沒有大量大型脊椎動物被取代。滅絕的物種包括猛獁象、乳齒象、地懶和巨型海狸,統稱為巨型動物群。晚更新世大型動物的滅絕不是同步發生的,也不是全世界同等規模的。
對於不同大陸上大型動物滅絕的時間存在相當大的疑問。目前,有證據表明,最早的大規模物種滅絕發生在澳大利亞和新幾內亞大約3萬年前。在這段時間內,大型有袋類動物如雙齒龍、爬行動物如7公尺長的goanna、Megalania和大型的不會飛的鳥類都消失了。澳大利亞脊椎動物屬中86%的成員體重超過40公斤的物種也滅絕。
早在更新世早期,非洲、亞洲和歐洲發生了小得多的滅絕事件,大得多的物種如犀牛、大象和最大的偶蹄類動物都消失了。大約12000年前,其他大型動物群滅絕事件發生在歐亞苔原(影響猛獁象、愛爾蘭麋鹿和毛茸茸的犀牛);在北美洲和南美洲,它們發生在大約11000年前(影響到各種各樣的物種,包括象、巨樹懶、獅子和熊)。這些物種的滅絕使歐洲29%的體重超過40公斤的脊椎動物屬和北美的73%的脊椎動物屬滅絕。
直到1000年至2000年前,紐西蘭和馬達加斯加等大而與世隔絕的大陸上的巨型動物群才得以生存。在過去的幾千年裡,馬達加斯加的象鳥和紐西蘭的moas等巨型鳥類在更新世之後消失了。
晚更新世滅絕的原因仍然存在爭議;然而,人們普遍認為,人類的到來預示著紐西蘭和馬達加斯加最近的滅絕事件。早期的事件還不太清楚,研究人員在人類引起的變化還是氣候變化是主要原因之間存在分歧。
最近,大型動物滅絕對植被和氣候的影響,特別是在澳大利亞,受到了人們的關注。澳大利亞大型動物群滅絕,火災發生率增加,可能導致植被結構變化,導致有效降水量減少,土壤更加貧瘠,甚至在其他有利條件下,愛爾湖無法填滿。
在過去的一萬年裡,生物圈發生了巨大的變化。農業和畜牧業的發明,以及這些做法最終在全世界的傳播,使人類能夠選擇地球上可利用生產力的很大一部分。計算表明,人類目前使用的太陽能量約占陸地上有機體捕獲的太陽能量的40%。在地球歷史上,一個動物物種利用地球上如此大比例的生產力是獨一無二的。
人口繼續以每年約8000萬人的速度增長,在21世紀的某個時候可能會達到100億。複雜的技術和不斷增長的人口造成的大氣變化有可能對生物圈造成重大破壞。其中最重要的變化是向大氣中排放包括二氧化碳、甲烷和氯氟烴在內的溫室氣體。
儘管在過去的幾十年裡,在理解生物圈方面取得了巨大的進步,但顯然還有很多東西需要學習。許多人會同意,我們才剛剛開始認識到維持生物圈適宜生命的複雜過程。