

第三章 生物群落和生態系統
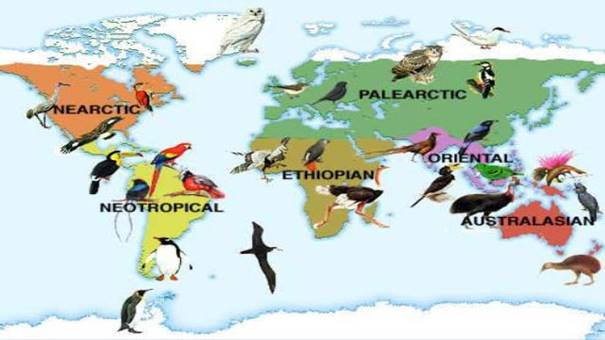
地球上最大的生態群落是生物群落biomes和生物地理區域。每一個巨大的系統都有一套獨特的物種和氣候條件。然而,顯而易見的模式出現了。世界各地的沙漠具有高氣壓和低濕度的特點,而不同大陸的熱帶雨林則有充足的水分和溫暖的溫度。這些氣候條件促成了每個地區的主要植被類型。(沙漠的特點是稀疏的灌木植被,而熱帶森林則由豐富多樣的厚冠層高大樹木組成。)本章致力於描述不同生物群落和生物地理區域的獨特特徵,以及某些生物群落與其他生物地理區的共同特徵。
生物地理區是動物和植物分佈的一個區域,具有相似或共同的特徵。根據一般經驗,陸地和內陸水域的動植物在世界上的一個地區到另一個地區或多或少都有不同。為什麼要這樣?為什麼在適合它們的環境條件佔優勢的地方就不存在同樣的物種呢?
世界各地具有相似環境條件的地理區域能夠容納相同類型的生物群。這種情況有效地將生物圈劃分為具有相同氣候條件和地質特徵、支援具有相似生命策略和適應能力的物種的生物群落。生物群落通常被描述為具有相似生命形式和環境條件的主要動植物群落。它包括各種各樣的群落,以主要的植被類型命名,如草原或針葉林。幾個相似的生物群落構成了一個生物群落類型,例如溫帶落葉林生物群落類型包括亞洲、歐洲和北美的落葉林生物群落。“主要生活區”是歐洲對北美生物群落概念的說法。
生物群落是較大的生物地理區域(植物界和動物界)的基本單位。熱帶森林是陸地生物群落的一種類型。它位於地球周圍的不同地點,那裡的氣候和地質條件產生了相似的環境。熱帶森林生物群落在任何地方都包含相同的生物群落。然而,從一個熱帶森林到另一個熱帶森林,個體物種將不一樣。取而代之的是,每一片森林將支持生態上相當的生物,即具有相似生命週期和類似於環境條件的不同物種。
動植物在各種生物群落中的獨特分佈是如何形成的,不能單純通過目前的氣候因素和緯向分帶來解釋。地質事件,如大陸漂移和過去的氣候條件也必須考慮在內。這是歷史生物地理學用來研究世界各地動植物分佈的方法。
生物地理學的概念

植物收藏家和系統學家約瑟夫·道爾頓·胡克爵士(Sir Joseph Dalton Hooker)的植物地理學工作和阿爾弗雷德·羅素·華萊士(Alfred Russel Wallace)的動物地理學工作對查理斯達爾文的工作產生了很大的影響。因此,達爾文進化論深深植根於對這個時代新興的生物地理學的理解。在《物種起源》一書中,達爾文包括了地理分佈的兩個關鍵章節,其中他提到了胡克和華萊士。在熱帶的高海拔地區,胡克發現了通常局限於溫帶地區的植物,達爾文將這些觀察結果解釋為過去氣候變化的證據。達爾文還採納了華萊士關於島嶼間動物群分佈的觀點:那些有相似動物群的島嶼只有淺水區分開,而且曾經是一塊毗連的陸地,對動物的擴散沒有阻礙,而那些動物群不同的島嶼被一直存在並阻止物種遷徙的深海航道隔開。

生物分佈
地理因素在分類學的各個層次都起著重要的作用。由於地理屏障而變得孤立的種群將趨向於與其物種的分化。雖然這些屏障包括海路、河流、山脈、沙漠和其他惡劣環境,看起來很小,但它們卻會在分類群之間造成一個楔子,最終導致相關物種、屬、科等(在分類層級上)發生分化。這種機制的一個例子出現在Gregory裂谷,東非裂谷系的東部分支;不同的牛羚亞種分佈在裂谷兩側,亞種產於東側和西側。其他哺乳動物,如斑紋角馬或王冠猴(cercopitechus mitis)也表現出類似的地理變異。非洲中部寬闊的剛果河是許多靈長類同類物種(同一屬的物種)之間的屏障,例如在剛果河北側發現的普通黑猩猩(Pan troglodytes)和生活在河以南的侏儒黑猩猩(P.paniscus)或倭黑猩猩(bonobo)。生活在不同大陸的同一科的屬之間有更重要的生物地理劃分,非洲象(Loxodonta)和亞洲象(Elephas)就是如此。整個科或亞目可能因生物地理領域的不同而有所不同,如在非洲和亞洲發現的舊大陸猴子(卡塔林猴)和南美洲的新大陸猴(platyrrhines)的靈長類分類中可以看到。
擴散論與生物地理學
在歷史生物地理學中,生物分佈模式的擴散論和生態變異假說一直存在分歧。根據擴散清單的觀點,物種形成發生在動物從起源中心向外擴散時,它們跨越了先前存在的屏障,而這些障礙是它們不容易重新跨越的,並且會切斷它們與原始種群的聯繫。生態變異的解釋指出,一個物種,在一個廣闊的區域內,隨著屏障的發展而變得支離破碎,就像在大陸漂移過程中發生的那樣。然而,這些模式並不是相互排斥的,它們都提供了對生物地理分佈模式的洞察。傳統的生物地理學家和主要的動物地理學家,如William Diller Matthew、George Gaylord
Simpson和Philip J. Darlington, Jr.—接受了許多關於物種分佈和分化模式的解釋,這些解釋通常屬於分散論觀點。
在20世紀50年代和60年代的一系列作品中,特立獨行的委內瑞拉植物地理學家Leon Croizat強烈反對這種物種分佈的分散列表解釋,他將其解釋為用於解釋生物地理分佈的特殊事件。他堅持認為,生物地理關係中的規律性太大了,不能用偶然跨越障礙來解釋。20世紀70年代,他的作品引發了維卡裡亞主義理論的發展。
儘管這些觀點在生物地理學中存在兩極分化,但分佈模式可以用分散清單和生態變異生物地理學相結合來解釋。許多生物地理學者認為,生態變異過程形成了分佈多樣性的基本機制,分散清單模式的運作更為零散。
地方性
分佈在某一特定地區的一個分類群被稱為該地區特有種。分類群可以是任何級別的,儘管它通常在科或以下,其分佈範圍可能很廣,橫跨整個大陸,或者非常狹窄,僅覆蓋幾平方公尺:一種松鼠(Sciurus kaibabensis)是亞利桑那州凱巴布高原的特有物種,靈長目狐猴科是馬達加斯加特有的,哺乳動物亞綱原喉亞綱(單孔目)則是澳大利亞的特有物種。新特有種(低等級分類群[例如物種]沒有時間擴展到其起源地區以外)和古特有種paleoendemics(尚未滅絕的高等級分類群[如階級]。
地方性的概念很重要,因為過去生物地理區域的制定是基於它。一個區域的界限是通過繪製分類群的分佈圖來確定的;在許多分類群的外部邊界出現的地方,劃定一個生物地理區域的界限。主要地區(領域和領域)仍然被確定為那些擁有最獨特性的地區,或者換句話說,那些與其他地區共用最少分類群的地區。隨著區域進一步細分,它們將包含更少的獨特分類群。
這種方法一直受到批評,因為它假設物種範圍是穩定的,而事實並非如此。確定生物地理區域的另一種方法是計算地理區域之間的相似程度。區域的相似性可以用Jaccard的生物相似性係數來量化,該係數由以下等式確定:

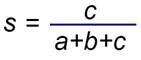
大陸橋
陸橋是在不同時期連接地球主要陸地的幾個地峽中的任何一個,其結果是許多動植物物種將它們的活動範圍擴展到新的地區。在古近紀、新近紀和第四紀的大部分時期(大約開始於6550萬年前),一座對新大陸動物群產生深遠影響的陸橋從西伯利亞延伸到阿拉斯加,但也有一些中斷。在這片狹長的土地上,有許多起源於舊大陸的生物,包括智人。
另一個重要的陸橋,巴拿馬地峽,在古近紀和新近紀的大部分時間裡被淹沒,結果北美和南美的動物群基本上是分開進化的,除了上新世時期(大約530萬年前到260萬年前)有幾十萬年的時期,當地峽被抬高的時候。
物種多樣性組成
物種多樣性不僅取決於生物群落中物種的數量,即物種豐富度,還取決於該群落中個體的相對豐度。物種豐度是指每個物種的個體數量,相對豐度是指群落中物種間個體分佈的均勻性。兩個群落的物種可能同樣豐富,但相對豐度不同。例如,每個群落可能包含5個物種和300個個體,但在一個群落中,所有物種都是相同的(例如,每個物種有60個個體),而在第二個群落中,一個物種的數量遠遠超過其他4個物種。
物種多樣性的這些組成部分對不同的環境條件有不同的反應。一個沒有種類繁多的棲息地的區域通常是物種貧乏的;然而,能夠佔據該區域的少數物種可能很豐富,因為與其他物種的資源競爭將減少。
物種豐富度的趨勢可以很好地揭示一個地區過去和現在的狀況。南極大陸的物種很少,因為它的環境太不適宜居住;然而,海洋島嶼物種貧乏,因為它們很難到達,或者像印尼中南部的小巽他群島那樣,因為它們的起源相當近,生物沒有足夠的時間來建立自己。
全球梯度也影響物種豐富度。最明顯的梯度是緯向的:熱帶的物種比溫帶或極地的物種多。生態因素通常被用來解釋這種等級。
更高的溫度、更大的氣候可預測性和更長的生長季節共同創造了一個更吸引人的棲息地,允許更多的物種多樣性。熱帶雨林是最豐富的棲息地,熱帶草原比溫帶草原表現出更多的多樣性,熱帶或亞熱帶地區的沙漠比溫帶沙漠擁有更廣泛的物種。
另一個影響特定區域物種豐富度的因素是將該區域與潛在物種來源隔開的距離或屏障。物種到達遙遠的海洋島嶼或孤立的山谷的可能性很小。動物物種,特別是那些不飛的物種,比植物物種更不可能這樣做。小巽他群島的氣候和植被與東爪哇島相似,但嚴格意義上的陸生動物卻少得多。這種情況是由於這樣一個事實,即Java在過去曾與較大的陸地相連,而較小的巽他群島則沒有。當植物和種子被風吹過中間的海洋時,幾乎沒有沒有沒有翅膀的動物物種到達這些島嶼。

物種對環境變化的適應
環境和有機體都不是靜態的。因此,兩者中任何一個的變化都會破壞兩者之間的關係。一個有機體的微小變化實際上可以改善相互作用—一個隨機的基因突變允許植物利用一種已經存在但以前不能被植物利用的營養物質,將提高有機體的生存能力。然而,極端性質的變化幾乎總是不適應的。微小的環境變化可能會帶來一個挑戰,生物體可以通過增加生理反應來應對,或者,如果它們是可移動的,則可以將自己轉移到壓力較小的區域。然而,災難性的破壞可能會造成一個不再適合這些有機體的環境,它們可能因此而消亡。
雖然物種的分佈模式是由環境條件決定的,但一個物種的實際範圍與它的潛在範圍並不相同,即在生態上與其需要相適應的區域。例如,世界上的生物地理區域與氣候因素有關,但它們並不與氣候因素共存。因此,位於北緯30°和南緯30°的沙漠生物群落和赤道附近的熱帶雨林生物群落,可以在大多數植物地理領域和動物地理領域找到。
地質變化對生物分佈的影響
20世紀60年代形成的板塊構造理論,現在已經牢固確立。它對大陸塊動態性質的解釋不僅在地質學領域,而且在生物地理學領域都很重要;它徹底改變了對動植物分佈的解釋。大陸的緩慢移動被用來解釋人口的孤立和混雜。在接受這一觀點之前,大陸橋和沉陷大陸被認為是過去地質學中大陸聯繫的手段。雖然大陸橋,例如連接北美西部和亞洲的白令海峽陸橋,已經存在並促進了生物的擴散,但人們認為它們不再像以前認為的那樣無處不在,在這一過程中發揮了重要作用。像Archelenis這樣假設的陸橋,據說連接南美和非洲西南部,現在被大多數專家視為早期傳記作家豐富想像力的遺跡。
在中生代的大部分時間(2.51億至6550萬年前),大陸形成了一個被稱為盤古大陸的整體。在白堊紀早期(1.455億至9960萬年前),特提斯海道形成並將盤古大陸分為北部大陸蘿拉西亞(包括歐亞大陸和北美)和南部大陸岡瓦納大陸(包括南美洲、南極洲、非洲、印度和澳大利亞)。儘管有短暫和移動的陸表海道,動植物基本上能夠在南北半球內自由移動,但不能在它們之間自由移動。在晚白堊世和整個新生代的大部分時間裡,岡瓦納大陸分裂,其組成部分漂移,其中一些與蘿拉西亞形成聯繫,後者或多或少保持著連續的陸塊。根據這一模型,澳大利亞自始新世以來(5580萬至3390萬年前)一直與其他大陸保持分離,從晚白堊世開始,澳大利亞只與已經極地的南極洲接觸,這有助於解釋其顯著不同的動植物群。南美洲的生命形式只不過不如澳大利亞獨特。自始新世以來,南美洲與其他大陸分離,直到上新世(530萬至260萬年前)才與北美建立永久聯繫。只有在那時,一些交換,特別是動物群,才被允許。非洲在古新世時期(6550萬至5580萬年前)已接近蘿拉西亞,此後與歐亞大陸的聯繫一直很薄弱,因此其目前的動植物群與東半球其他熱帶地區更為相似。印度在古近紀時期與蘿拉西亞有著廣泛的聯繫,因此沒有明顯的(古特有的)生物。
動植物分佈邊界

把動植物分類到主要的生物地理區域是一項有用的、產生假說的活動。當兩個生物類群在分佈上表現出相似的變化時,理論上認為它們受到了相同種類的進化過程的影響,例如有利於某些適應或隨機地理變化的生態限制。在對一個生物群落中許多分類群的調查中,它們可能都有相似的分佈模式;它們可能受到相同地理屏障的限制,也可能受到氣候因素的類似影響。當比較植物地理領域和動物地理領域時,人們會被輪廓上的廣泛一致和細節上的差異所打動。
在這些模式中確實存在奇怪的差異。一些生物能夠“跳過”氣候帶,因此它們在北溫帶和南溫帶都能找到,但在中間的熱帶地區卻找不到。其他人似乎有著特殊的能力,能夠分散到偏遠、孤立的地區並生存下來。例如,鳥科的Rallidae(rail)成員分散在許多島嶼上,包括新赫里多尼亞島、豪勳爵島、關島,甚至被恰當地命名為不可接近的島嶼,在南美洲西海岸的加拉帕戈斯群島和非洲東海岸的塞舌耳群島都發現了這種巨型烏龜。
動植物分佈也存在差異。例如,一個獨立的領域,南非(Capensic)領域,被認為是植物而不是動物。在新幾內亞,植物區系被歸為古熱帶區系,但動物群不屬於相應的古熱帶區系,而是被歸為Notogaean區系。其中有些差異比其他的更容易理解。動物區系的缺乏可能僅僅是動物活動性增強的結果。這種劃分,如果真的存在於動物地理學中的話,已經被周圍的新大陸和非洲熱帶動物群“吞沒”。其他的差異,特別是新幾內亞動植物的差異,不太容易解釋。
陸地和淡水植物群比它們共存的動物群更古老;因此,主要的植物地理區域反映了地球歷史上比動物地理區域更古老的階段。因為植物的活動性較低,它們的聯合體一直存活到現在相對完整。將主要區域劃分為幾個小的分區有助於闡明地球歷史上最近發生的事件,以及生物群的擴散能力、適應策略和生態關係。
氣候對生物擴散影響的重要性不容忽視。海洋生物往往沿氣候線分佈,許多陸生類群,如候鳥,由於活動性強,已經分佈在兩個或兩個以上的主要生物地理區域。儘管它們分佈廣泛,但它們在南北溫帶地區分佈很廣,它們之間被不適宜的熱帶地區隔開。
這些奇怪的、不連續的分佈提醒我們,生物地理區域只勾勒出有機分佈的輪廓,它們並不能解釋每一種情況。它們的用處在於指出擴散機制、過去的氣候走廊和其他重要的生物現象。
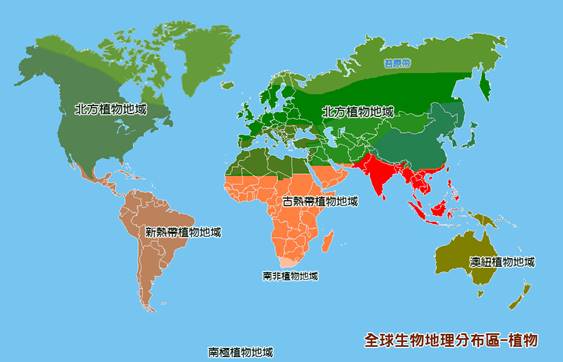
植物地域分布 F loral K ingdoms
植物分布六個大地域,包含:北方域Boreal(全北極Holarctic))、古熱帶域Paleotropical、新熱帶域Neotropical、南非開普敦域(Capensic)、澳大利亞域和南極域共六個植物地域。
這些地域被進一步細分為副地域和地方區域,對此存在一些爭議。植物界的劃分並不明顯,高等植物的科在植物地理界的分佈程度也各不相同,它們的分佈僅部分取決於它們的年齡。以下分類是根據隆納德(Ronald Good)1974年的研究。
北方植物地域分佈 Neotropical Kingdom
北半球地域(Neotropical kingdom)由歐亞大陸和北美洲組成,自始新世(5580萬至3390萬年前)以來,這兩個地區基本上是一個毗連的地塊。西伯利亞和阿拉斯加之間狹窄的白令海峽,只存在于更新世末期(約11700年前)。這兩個大陸植物區系的差異很小,這一點也不奇怪。樺木科、芸苔科、報春花科、虎耳草科、薔薇科、毛茛科、傘形科等科分佈在北半球的溫帶地區。北半球地域分為六個副地域:
(1)北極和亞北極地區是北方苔原帶,從斯匹茨伯根(北冰洋中的一個島嶼,在挪威以北)沿著北冰洋沿岸延伸,經過西伯利亞和北美洲北冰洋,一直延伸到格陵蘭島。該地區的開花植物多樣性較差,但隱花植物更為多樣。
(2)東亞或中日地區有大約300個特有屬,從東喜馬拉雅山脈的斜坡延伸到中國東北部和俄羅斯遠東地區,包括臺灣、日本和薩哈林島。在這個地區,南部的熱帶雨林與北部的落葉林合併。典型的植物科有月桂科、木蘭科和茶科。有許多特有屬;小檗屬、杜鵑花屬和刺柏屬是山區特有的屬。
(3)以中亞和蒙古沙漠草原為中心,西亞和中亞地區是一個由200多個特有屬組成的植物區系。它從高加索延伸到西藏高原,分佈著藜科(鵝掌)的乾旱地帶植物和柳屬(柳樹)、黃芪(奶紫雲杉)和雲杉(雲杉)。
(4)地中海地區是北極領域的冬季降雨區。以葉狀植物為主要特徵的叢生型。但很難界定,因為它的許多特色植物(約250屬)都集中在這一地區,但並不局限於這一地區。該地區完全沿著地中海延伸,從葡萄牙到敘利亞。一些分類將包含亞熱帶雨林生物群落的加那利群島放在這一地區,但Good將這些島嶼與其他東大西洋島嶼群歸入一個單獨的馬卡羅尼西亞地區,該地區約有30個特有屬。
(5)歐洲西伯利亞地區從冰島經西伯利亞延伸到堪察加半島。松科松樹(松樹)、落葉松(落葉松)、雲杉(雲杉)和冷杉(冷杉)科的針葉樹生長在廣闊的單株林分中,南部為溫帶落葉林,北部為苔原,沼澤地(在適當的地區包含杜鵑花科[石南科]、苔草[莎草]和泥炭蘚)。該地區西部的物種比東部豐富得多:大約有100個屬是歐洲特有的,只有12個屬是西伯利亞特有的。
(6)白令海峽以東的北美地區的植被與西部歐洲西伯利亞地區的植被非常相似,只是略有不同。針葉樹屬鐵杉屬(鐵杉屬)、紅杉屬(紅木屬)取代了歐洲西伯利亞的同類植物,有9個特有的開花植物科。Good和其他分類群將北美洲的東(大西洋)和西(太平洋)分為不同的區域,大西洋地區特有100屬,太平洋特有300屬,儘管這些特有分類群只占總植物區系的一小部分。
古熱帶植物地域及其副地域 Paleotropical Kingdom
這個植物地域從非洲延伸,不包括南北邊緣地帶,穿過阿拉伯半島、印度和東南亞,向東進入太平洋。延伸到該地區大部分地區的植物科包括Pandanaceae(螺旋松)和Nepenthaceae(東印度豬籠草)。然而,這一巨大地區的植物群並不同質:98%的夏威夷植物群物種是特有的,斐濟70%的花卉物種和新赫里多尼亞60%的花卉物種也是地方性的。領域的劃分是有爭議的,但最常見的是馬列西亞,印度和波利尼西亞亞領域。
馬列西亞亞副地域包括東南亞島嶼和馬來半島,向東延伸到新幾內亞大陸。雖然有時它也被印度歸入印度-馬來語地區,但C.G.G.J.van Steenis(1950)所稱的馬來西亞植物群形成了一個緊密的統一體,可細分為三個部分:西部地區,包括馬來半島、蘇門答臘、婆羅洲和菲律賓;爪哇島和小聖代的南部地區;以及西裡伯斯、摩鹿加群島和新幾內亞的東部地區。該地區約有400個特有屬(占地球植物區系總數的20%),其中西區130屬,南區15屬,東區150屬。生物群落類型從熱帶雨林到山地和雲林,南部地區的生物群落類型較為乾燥。該地區西部的熱帶雨林生物群落的特點是以龍腦香科為主,儘管在整個地域也發現了杜仲科、桑科和番荔枝科。
在印非副地域,包含非洲植物群(撒哈拉以南)和印度次大陸、緬甸和中國南部幾乎沒有區別。這些地區由一條貫穿阿拉伯半島和伊朗南部的走廊緊密相連。馬達加斯加島的植物區系在該地區最為不同,通常被視為形成一個獨立的地域;該島有12個特有科和350個特有屬,儘管它們只占總數的四分之一左右。斯里蘭卡的植物區系與馬列西亞亞副地域幾乎和印度有著同樣多的共同點。植被範圍從雨林到半乾旱草原。豆科(豆科)和紫菀科(紫菀科),通常被稱為菊科,在該地區擁有最大的多樣性,與山茱萸科(印度杏仁)以及在馬達加斯加南部乾旱地區的Didiereceae一起。大草屬和巴本松屬(包括大草屬和巴本森屬)。在山地副地域(非洲高山區),須芒草屬、柳枝稷和猴麵包樹是其特色。大約50個特有屬界定了從撒哈拉沙漠到印度西北部的沙漠地帶;500個在熱帶非洲特有,120個在印度,300個在東南亞大陸,但這些地區的邊界界定不明確,特有種的分佈也只是微弱的共有。
在許多方面,太平洋島嶼是馬列西亞的異類,但波利尼西亞亞領域夏威夷、玻利尼西亞其餘部分、美拉尼西亞和密克羅尼西亞、新赫里多尼亞以及豪勳爵和諾福克群島的四個主要分區中,每一個都有大量的地方性分類群。夏威夷有40多個特有屬;波利尼西亞(不包括夏威夷)有近20個屬;美拉尼西亞和密克羅尼西亞地區有38個,其中17個僅限於斐濟;新赫里多尼亞有135個,島上共有600個屬。只有21個亞領域特有屬出現在四個分區中的一個以上。植物區系的不平衡表現在檳榔科(有時稱為棕櫚科)特有種中的優勢。在波利尼西亞亞領域和其他一些科中,棕櫚科有35個以上的特有屬。
新熱帶植物地域 Neotropical Kingdom
從本質上講,新熱帶領域覆蓋了除南美洲最南端和西南地帶以外的所有地區;中美洲;墨西哥,不包括乾燥的北部和中部;以及西印度群島和佛羅里達州南端。植被範圍從亞馬遜河和奧里諾科盆地的熱帶雨林到委內瑞拉(利亞諾斯)和阿根廷(潘帕斯)的開闊草原。有47科近3000屬的開花植物是這個領域特有的;有些科,包括鳳梨科(鳳梨)和仙人掌科(仙人掌),實際上都局限在這個領域。在領域內部,中美洲,包括墨西哥和地峽、西印度群島、委內瑞拉-圭亞那地區、巴西、安第斯山脈和潘帕斯地區都有一定程度的地方性。雖然貧窮,胡安費爾南德斯群島和德斯文圖拉多斯群島,位於智利西海岸,顯示出高度的地方性和普遍的新熱帶親和力。
南非植物地域 South African Kingdom
南非,或稱卡彭斯地域(Capensic),由非洲的南端和西南端組成,即好望角周圍的地區(因此得名為“卡彭斯”)。它的植物非常豐富,有11科500屬。這是最小的植物地理領域。冬季降雨氣候模式與地中海地區相似,植被的總體面貌與該地區的灌木叢植被(maquis)相似。在這個狹小的限制區的邊緣,植物群與非洲古熱帶的典型植物群融合在一起。
澳 紐 植物地域 Australian Kingdom
澳洲大陸形成了一個與古熱帶截然不同的領域。熱帶雨林生物群落從北部的熱帶雨林到南部的溫帶雨林,特別是塔斯馬尼亞的熱帶雨林,出現在東海岸。桉樹林地覆蓋了該大陸東部三分之一的大部分地區,在西南部發現了由顯著的溫帶森林和班克西亞荒原組成的馬賽克。(澳大利亞植物區系的這兩個元素雖然引人注目,但不是地方性的;在新幾內亞東部、新不列顛、小巽他和菲律賓有一些桉樹,在新幾內亞發現了一種Banksia。)否則,大部分植被是半乾旱或適應乾旱的。約有19科500屬為特有種。只有昆士蘭東北部的熱帶雨林才有混合植物群,帶有顯著的馬來元素。
南極植物地域 Antarctic Kingdom
這個植物地域包括南美洲的南端,沿著智利海岸向北延伸一段距離;紐西蘭;以及南極和亞南極島嶼。在紐西蘭南島、塔斯馬尼亞島和澳大利亞阿爾卑斯山,南極和古熱帶植物群以一種有趣的交錯模式出現。根據古德所說,在這個領域裡大約有50個屬是常見的。
智利南部、巴塔哥尼亞和紐西蘭組成亞南極地區。它有一個獨特的森林植物群,其中沒有山毛櫸(南方山毛櫸)可能是最具特色的元素。
南極地區包括南極島嶼和大陸邊緣地區。這個地區的植物非常貧乏。一般來說,開花植物不能很好地適應惡劣的氣候,而苔蘚和其他隱花植物是主要的組成部分。在紐西蘭和澳大利亞南部,尤其是塔斯馬尼亞,海拔較高的地方可以找到真正南極植物群的蹤跡。
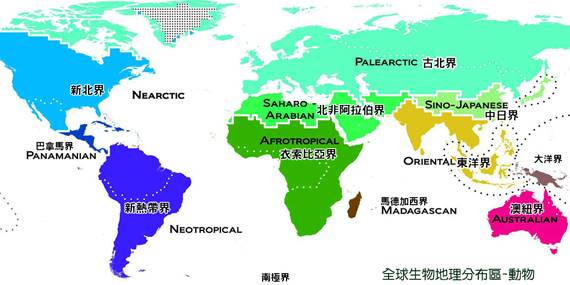
動物領域分布 F aunal R ealms
儘管對動物地理分佈的最早研究是1858年對斯萊特的研究,但在他的經典著作《動物地理分佈》(1876年)中,是華萊士設定了確定動物地理區域或領域的參數。華萊士發現了三個領域:Megagaea或Arcotogaea,包括非洲、歐亞大陸和北美;Notogaea,包括澳大利亞、大洋洲和紐西蘭;Neogaean,包括中美洲和南美。他的劃分雖然有所改變,但構成了今天公認的領域的基礎。
儘管不同物種具有不同的擴散能力,但傳統的動物地理邊界可以解釋鳥類和昆蟲的分佈。總的來說,陸生哺乳動物、淡水魚和無脊椎動物的分佈似乎很一致,並提供了動物地理分區的最佳證據。
在許多情況下,動物群混合的區域已經得到了很好的研究。有些分類任意地將它們包含在一個區域(或領域),有些分類在任何形式的賦值中忽略它們,並將它們放在減法轉換區。這種區域的一個例子是華萊士,它包括菲律賓、西裡伯斯、摩鹿加群島和小巽他群島。位於古熱帶和澳大利亞之間,華萊士包含兩個地區的混合體。動物群貧瘠且不平衡,但該地區具有很高的地方性。
以下部分是基於P.J.達林頓的研究,並在很大程度上修改了這些部分。
華萊士線 Wallace’s Line
華萊士線是19世紀英國博物學家阿爾弗雷德·羅素·華萊士提出的東方和澳大利亞動物區界線的名稱。該線路從印度洋穿過龍目海峽(巴厘島和龍目島之間),向北穿過馬卡薩海峽(婆羅洲和西裡伯斯之間),向東,在棉蘭老島南部,進入菲律賓海。儘管許多動物地理學家不再認為華萊士線是區域邊界,但它確實代表了許多主要動物群分佈的突然極限。許多魚類、鳥類和哺乳類動物在華萊士線的一側有大量的代表性,但在另一側卻很少或根本沒有。
全北極動物領域 Holarctic Realm
全北極領域通常根據陸地生物分為兩個副領域:近北極領域(北美洲)和古北極領域(歐亞大陸和北非)。與北美植物地理分區不同,近北極區的動物地理區向南延伸,包括整個佛羅里達州和下加利福尼亞。在全北極領域發現了一些有趣的間斷分佈:一些分類群在歐洲和北美洲東部之間,一些在歐洲和東亞之間,另一些在北美西部和東亞之間。這些分佈也許可以根據最近氣候帶的運動來進行解釋。
淡水魚和無脊椎動物的專家們喜歡把全北極領域劃分得更細。Petru Banarescu提出以下副領域劃分:(1)歐洲至地中海地區;(2)西伯利亞、貝加爾湖和蒙古西部;(3)北美東部、西部和北極地區;以及(4)墨西哥中部。
這一領域特有的科包括哺乳動物,如鼴鼠科、河狸屬、鼠兔科;兩棲動物,如蠑螈科、蠑螈科、隱鰓鯢科和洞螈科;以及無脊椎動物,如淡水小龍蝦科及螯蝦科。
古熱帶動物領域 Paleotropical Realm
古熱帶或非洲特提斯地區被清楚地分為兩個動物領域,這兩個區域有時被視為單獨的領域:非洲熱帶地區,包括撒哈拉以南和阿拉伯西南部的非洲大陸;東方地區,包括熱帶南亞和東南亞,包括相關的大陸島嶼。另外兩個地區,馬達加斯加和華萊士,通常與這兩個主要地區分開。
由於地理上的連續聯繫,古熱帶動物領域和全北極動物領域相互融合。然而,每一個都有許多不同的元素,部分但不完全是因為它們的氣候不同。哺乳動物石龍目(穿山甲)和長鼻目(大象)是古熱帶地區特有的。局限於該領域並延伸至該領域的哺乳動物家族包括:尾猴科(舊大陸猴子)、懶猴科(蜂猴、嬰猴、金熊猴和樹熊猴)、豪豬科(舊大陸豪豬)、靈貓科(麝貓和獴)、犀牛科(犀牛)和鼷鹿科(鼷鹿)。特有的鳥類科包括佛法僧目犀鳥科(犀鳥)和八色鶇科(八色鶇);以及特有的爬行動物科,變色龍科(舊大陸變色龍)。
非洲熱帶動物領域(或衣索比亞動物領域)和全北極動物領域之間的界線通常劃過撒哈拉沙漠。然而,查理斯史密斯(Charles H.Smith)對哺乳動物分佈的徹底再分析得出結論,地中海地區,包括其南部和北部海岸,實際上在古熱帶地區比全北極地區更具古熱帶特徵。嚴格地說,非洲特提斯海(指中生代將蘿拉西亞和岡瓦納大陸分隔開來的水道特提斯海)一詞適用於這一擴大的概念。
與構成南非卡彭斯領域的非洲南端的植物生活形成鮮明對比的是,開普敦地區的動物群與周圍地區的動物群無法區分。據推測,可能曾經存在過的任何獨特的動物區系卡彭斯Capensic元素都與熱帶元素融合在一起。非洲大陸特有的分類群包括哺乳動物蹄兔目(蹄兔)、土豚科(土豚)和象鼩目(象鼩);哺乳動物科金毛鼴科(金鼴)、跳兔目(跳兔springhares)、蔗鼠科(蔗鼠cane rats)和長頸鹿科(長頸鹿和okapi);鴕鳥科(鴕鳥), 鵜形目(鯨頭鸛)和鷺鷹科(蛇鷲);青蛙亞科姬蛙;淡水魚亞綱多鰭魚科bichirs、象鼻魚科(伯氏長頜魚snoutfish)和電鯰科malafteruridae(電鯰);蝸牛科陸地旋蝸牛。
馬達加斯加與非洲大陸大不相同,作為一個單獨的區域,它通常被給予平等的地位。與非洲大陸(古熱帶地區)共有的哺乳動物科包括馬島蝟科(馬島蝟Terrecs和巨型水獺)和河馬科(河馬最近在馬達加斯加滅絕)。馬達加斯加還與新熱帶地區有一些類群,特別是鬣蜥和蟒蛇,古熱帶地區的其他部分可能在古近紀Neogene和新近紀時期(6550萬至260萬年前)消失了。馬達加斯加特有物種包括哺乳動物中的幾個狐猴科。tenrec食蟲動物、食肉動物和鼠鼠齧齒動物的獨特亞群也很常見,象鳥科Aepyornithiedae(最近滅絕的象鳥)和其他鳥類亞科和科也是地方性的。熟悉的非洲大陸動物,如猴子、羚羊、大象、犀牛和大型貓科動物都不在馬達加斯加。塞舌耳Seychelles群島和馬斯卡林Mascarene群島有著遙遠的馬達加斯加親緣關係,一般都包括在馬達加斯加地區。

在東亞(或中印大陸)動物領域,特有的科包括,在哺乳動物中,樹鼩科(樹鼩)、跗科(眼鏡猴)和長臂猿科(長臂猿);在爬行動物中,有婆羅蜥科(無耳巨蜥)和長吻鱷科(恆河鱷);以及一些鳥類和無脊椎動物科。
河馬仍然在非洲大陸上遊蕩,但現在在馬達加斯加島附近已經滅絕,馬達加斯加島被認為是其獨特的生物地理區域
東亞大陸領域和非洲熱帶領域與全北極領域之間的三方邊界很難界定;基本上整個西南亞地區是過渡的。然而,這一範圍內的某些地區則更為明確:巴勒斯坦沙漠-內蓋夫Negev和阿拉伯紅海沿岸主要屬於非洲熱帶動物領域,而敘利亞、伊拉克、伊朗和阿富汗較少顯示出非洲熱帶動物領域特徵,並與全北極動物領域更多的聯繫。一種獨特的沙漠動物群,通常被稱為薩哈羅-辛蒂亞(Saharo-Sindian),將整個東北非洲與阿拉伯半島區涵括在一起,成為三個動物領域(非洲熱帶區、全北極區、中印大陸區)中間的過渡區域。
哺乳動物專家如G.B.Corbet將中國中部的東亞地區和全北極動物領域劃出了大致的邊界;然而,根據淡水魚類和無脊椎動物的證據,Banarescu將他所稱的中印大陸地區向北延伸,包括天山山脈系、西藏和黃河流域。
許多爭論都圍繞著東亞大陸動物領域和澳大利亞大南方(Notogaean)動物領域之間的分界線。華萊士認為亞洲大陸架(巽他大陸(棚)架)的邊緣構成了這一地區的邊界,華萊士的界線是婆羅洲、巴厘島和菲律賓以東的“典型”東方動物群的分界線。這種劃分的基礎是線的東部和西部的動物群之間的顯著差異。隨後幾代人一直在就這一邊界的位置進行辯論。該線的北部被赫胥黎T.H.Huxley改變,落在菲律賓西部(不包括巴拉望)。赫胥黎線被一些動物地理學家認為是一個更合適的劃定,因為菲律賓的動物群非常特殊。
著名的動物地理過渡區稱為華萊士線,位於印尼中部。這一地帶通常包括在古熱帶地區,西面以赫胥黎線為界,東面以沿澳大利亞大陸架(薩胡爾大陸架)邊界的萊德克爾線為界;它包括東亞和澳大利亞的動物群。韋伯線位於摩鹿加群島以西,代表著這兩種動物群均勻混合的地區。植物地域分界沒有明顯的。印尼西裡伯斯島Celebes和菲律賓群島,除了巴拉望島之外,都是屬於東亞動物領域,但也都有一些比較特別的動物群。這些動物群與它們的東亞近親不同,儘管有些動物群,如印尼西裡伯斯的動物群,也包含了一些澳大利亞元素。小巽他群島和帝汶地區一樣,擁有或擁有極少數但與眾不同的屬。在摩鹿加群島,動物群顯然與新幾內亞有密切關係。
大南方動物領域 Notogaean Realm
大南方動物領域,或大洋洲動物領域,從萊德克線(Lydekker’s Line)以東開始,延伸到太平洋。它由四個地區組成:澳大利亞、大洋洲、紐西蘭和夏威夷。然而,許多太平洋島嶼的動物群與古熱帶動物群和澳大利亞本土動物群有著相同之處。該地區特有的是單孔目動物(產卵哺乳動物,如鴨嘴獸)、六目中的四目、許多鳥類和魚類科以及一些無脊椎動物。岡瓦納人的親緣關係包括石鱉(不會飛的鳥)、肺魚、爬行動物科龜科(蛇頸龜)和已滅絕的小笠龜科(角龜)、青蛙科(樹蛙)和細足綱動物科,以及一些無脊椎動物科。
澳大利亞本土包括澳大利亞、新幾內亞和所羅門群島。鳥類目,如Rheoides(rheas)和Cashowaries(Casowaries)以及Menuridae(lyrebirds)和Paradisaeidae(天堂鳥)等科是該地區特有的,這是除了蝙蝠以外的所有哺乳動物的唯一一部分。將新幾內亞納入這一地區是很有趣的;新幾內亞動物群包括澳大利亞動物群的雨林部分。這個大島生物親和力的差異也許是植物地理學和動物地理學之間最顯著的差異之一。(如上所述,新幾內亞的植物群被歸類為古熱帶,但動物群不包括在可比較的動物區系範圍內。)
海洋區域界限不清。它包含了一些地方特有種,特別是新赫里多尼亞的鳥科Rhynochetidae(kagu)。許多動物群,尤其是鳥類,具有明顯的澳大利亞親緣關係。
紐西蘭地區包括整個紐西蘭,不包括西南部的動物群,西南部的動物群顯示出南極元素。不會飛的鳥棲息在紐西蘭和澳大利亞,儘管恐龍目(幾維鳥和摩亞鳥)是紐西蘭特有的。其他地方性分類群包括蝸牛科田螺科;紐西蘭唯一的哺乳動物蝙蝠科Mystacinidae;Xenicidae(紐西蘭鷦鷯);Leiopelmatidae(青蛙的一個原始科);和蝶骨科(tuatara,一個原始爬行動物科)。
夏威夷地區完全由夏威夷及其離島組成。該地區有一些特有的無脊椎動物科和一個鳥類科,即夏威夷蜜鼠科。
新區動物領域 Neogaean Realm
新大陸或新熱帶地區從墨西哥的熱帶低地向南延伸,穿過中美洲進入南美洲,一直延伸到溫帶和亞北極帶,包括西印度群島。在特有的哺乳動物類群中,自古新世(6550萬至5580萬年前)以來,就出現了雙足綱(有袋類動物的一個目)和幾個獨特的胎盤目,如無齒目動物(以及一些已滅絕的目)。到漸新世(3390萬到2300萬年前),鴨嘴獸(新世界的猴子)和一群齧齒動物(穴居動物)通過至今仍不為人所知的方式進入南美洲。在鳥類中,有兩個完整的目,即不會飛的雷形目(rheas)和䳍形目Tinamiformes(tinamous)以及30個科。一些魚類和無脊椎動物類群也都是地方性的。這些地方性分類群中的許多被認為可以追溯到岡瓦納時代(早白堊世),那時南部大陸形成了一塊大陸。這一觀點的證據來自於非洲和澳大利亞的近親—例如,平胸鳥類-不會飛的鳥、肺魚、蛇頸龜科、已滅絕的多角龜、樹蟾科、細趾蟾科和許多無脊椎動物(特別是原始的甲蟲,如天鵝絨蠕蟲)。
在西印度群島,這是一個新區動物領域中的特別地區(動物群稀少特別),獨特的哺乳動物包括兩個地方性的食蟲動物、溝齒鼩科和最近滅絕的島鼯。加拉帕戈斯群島也是一個動物群稀少特別的動物群,它們來自南美洲大陸,但卻獨立分化許多島嶼生態特徵。
南極動物領域 Antarctic Realm
南極動物領域,或稱為阿爾基諾Archinotic,包括南極大陸、南極鄰近島嶼和紐西蘭西南部分。該動物領域的存在,或者更確切地說是它的幽靈的存在,因為在今天的任何地方它都不存在於一個潮濕的狀態,這是由紐西蘭和南美洲常見的一些類群所證明的,如石蠅科Eustheniidae、甲殼綱海螺目和某些淡水螺類。有袋動物科微生物類群僅限於智利,與澳大利亞有袋類動物的親緣關係比其他南美有袋類動物更為密切,這一點似乎很有道理,它是南極聯繫的遺跡。
生態系統
幾乎所有生態系統的基本能源都是來自太陽的輻射能。太陽光的能量被生態系統的自養或自我維持的有機體所利用。這些生物主要由綠色植物組成,能夠進行光合作用,也就是說,它們可以利用太陽光的能量將二氧化碳和水轉化為簡單的、富含能量的碳水化合物。自養生物利用儲存在簡單碳水化合物中的能量來產生更複雜的有機化合物,如蛋白質、脂類和澱粉,維持生物體的生命過程。生態系統的自養部分通常被稱為生產者水準。
自養生物產生的有機物直接或間接維持著異養生物。異養生物是生態系統的消費者;它們不能自己製造食物。它們利用、重新排列並最終分解自養生物所形成的複雜有機物質。所有的動物和真菌都是異養生物,大多數細菌和許多其他微生物也是異養生物。
自養生物和異養生物共同構成了生態系統中不同的營養(攝食)水準。生產者層是由那些自己製造食物的有機體組成的。初級消費層由那些以生產者為食的有機體組成,第二消費層由以初級消費者為食的有機體組成,以此類推。有機物和能量從生產者層面到消費者層面的流動構成了一個食物鏈。例如,草原上典型的食物鏈可能是草(生產者)→老鼠(第一消費者)→蛇(第二消費者)→鷹(第三消費者)。
實際上,在許多情況下,生態系統的食物鏈相互重疊,相互連接,形成生態學家所說的食物網。所有食物鏈的最後一環是由分解者組成的,這些異養生物分解死亡的有機體和有機廢物。主要消費者以活植物為食的食物鏈被稱為放牧途徑;主要消費者以死植物為食的食物鏈被稱為碎屑途徑。這兩種途徑在計算生態系統的能量預算時都很重要。
生物群落
在決定群落整體結構的因素中,有物種(岐異性diversity)的數量、每種物種的數量(豐度abundance)、物種之間的相互作用以及群落在遭受破壞性影響(如火災或乾旱)後恢復正常的能力。生物群落隨時間的變化稱為演替或生態演替(succession)。
群落中的各種物種各自佔據著各自的生態位(niche)。一個物種的生態位包括它與群落其他成員的所有相互作用,包括競爭、捕食、寄生和互惠。一個群落中的有機體可以通過顯示哪個吃哪個而沿著食物鏈定位,這些位置被稱為營養級聯。第一個層次包括生產者,光合植物,它們將太陽的輻射能轉化為其他生物可利用的營養物質。
這些植物被食草動物(食草動物,或初級消費者)吃掉,這是第二個營養級聯。食草動物反過來又被食肉動物(肉食動物)吃掉,它們經常被較大的食肉動物(分別是第二和第三消費者)吃掉。當最後一個環節死亡並受到各種細菌和真菌的附著分解時,食物鏈就結束了,這些分解者分解了死去的有機物,從而將必要的營養物質釋放回環境中。
兩個不同植物群落之間的植被過渡區,森林和草原等兩種不同植物群落之間的植被過渡區被稱為交錯帶。它具有每個邊緣群落的一些特徵,並且通常包含重疊群落中沒有的物種。交錯帶可能存在于一個寬闊的地帶或一個小的口袋裡,比如森林空地,兩個當地社區融合在一起。兩個相鄰的群落相互之間的影響被稱為邊緣效應。一個生態區通常有更高的密度的一個物種和更多的物種數量比在任何一個側面的群落。有些生物需要一個過渡區域來進行求偶、築巢或覓食等活動。
