
第四章 陸地生態系統
地球在生命形式和物理特徵上都有巨大的多樣性。兩者都有助於生態系統的形成和發展。生態系統的結構決定並在某種程度上由生物參與者決定。取食和分解途徑以及物種之間和物種內部的競爭和其他相互作用的影響影響著生態系統中的營養循環和能量流動。儘管地球上存在著眾多的生態系統,下面描述的案例研究提供了生物、化學和地質力量如何相互作用,為生物創造獨特環境的樣本。
n 山地生態系統

喜馬拉雅-西藏地區是山地面積最大、最高的地區;最長的幾乎連續的山脈是沿美洲西海岸從阿拉斯加到南部智利的山脈。其他特別重要的山區包括歐洲(阿爾卑斯山、比利牛斯山脈)、亞洲(高加索、烏拉爾)、新幾內亞、紐西蘭和東非。
山地生態系統的起源
從地質時間框架來看,山地抬升和侵蝕的過程發生得比較快,因此高山山脈具有某種短暫的特徵。許多山脈與其他環境條件相似的地區隔離開來,它們的峰頂地區類似于新近形成的涼爽氣候島嶼,在大面積不同的溫暖氣候中定居下來。由於這種隔離,山頂上有一個獨特的生物群,由適應寒冷溫度的年輕動植物組成。然而,在低海拔地區,一些山脈能夠為環境變化所取代的更古老的生物群提供避難所。此外,山區植被通常比周圍地區受到人類活動的影響要小,因此可能藏匿著因其他地方發生的人為干擾而被趕出的動植物。
在過去200萬年的冰期期間,北半球適合於冷適應生物群的棲息地的冰期覆蓋了比現在大得多的區域,並且發生了大量適應冷的動植物的遷徙。北極生物群向南擴展,覆蓋了北美北部、歐洲和亞洲大部分地區的冰原。當氣候條件改善時,這些生物會向北退至北極緯度地區,並向山地地區進發。例如,這段歷史解釋了高山(如歐洲阿爾卑斯山和北極地區)動植物之間的密切相似性。
然而,在熱帶地區,由於熱帶低地的大片林地構成了遷徙的障礙,因此幾乎沒有機會在陸地上進行類似的冷適應生物活動。因此,這些有機體與其他寒冷環境中的有機體更加完全地隔離開來。儘管如此,熱帶高山的殖民活動還是發生了。鳥類的活動性特別強,一些溫帶的親緣關係可以到達赤道的山峰;例如,在新幾內亞的山區,可以發現在鄰近的熱帶低地沒有近親的琵琶和畫眉。候鳥可能是在同一地區生長的冷適應植物種子的載體,而這些地方也缺乏熱帶低地的親緣關係。
山地物種的種群通常都很小,雖然波動很大,而且是孤立的,而且往往是在相對較短的時間內進化而來的。因此,在不同的山峰上遇到相關但不同的物種並不罕見。這種新物種的迅速繁殖極大地促進了山地的生物多樣性和生物重要性。
生物多樣性,也叫生物多樣性,是指在地球上某個地方發現的生命的多樣性,或者,通常是地球上生命的全部多樣性。衡量這種多樣性的一種常用方法,稱為物種豐富度,是一個地區的物種數量。例如,哥倫比亞和肯雅各有1000多種鳥類繁殖品種,而大不列顛和北美洲東部的森林卻不到200種。北部的珊瑚礁
澳大利亞可能有500種魚類,而日本岩石海岸線可能只有100種。這些數位反映了熱帶地區之間的一些差異,例如,熱帶地區比溫帶地區擁有更多的生物多樣性,但原始物種數量並不是多樣性的唯一衡量標準。此外,生物多樣性包括每個物種的遺傳多樣性和物種所創造的生態系統的多樣性。
衡量物種多樣性的第二種方法是認識到那些只有少數物種但不常見物種的環境的獨特生物多樣性。引人注目的例子來自極端環境,如南極活火山(如羅斯海地區的埃雷伯斯山和墨爾本山)、溫泉(如美國西部的黃石國家公園)或深海熱液噴口。在這些地方發現的物種數量可能比其他任何地方都要少,但這些物種是非常獨特的。
除了物種間的多樣性,生物多樣性的概念還包括物種內部的遺傳多樣性。一個例子是人類物種,因為人類在許多方面的特徵上存在差異,這些特徵部分或全部是由遺傳決定的,包括身高、體重、皮膚和眼睛的顏色、行為特徵以及對各種疾病的抵抗力。
地球生物多樣性的目錄是不完整的。大約150萬種有學名。據估計,現存物種總數約為1000萬種,這意味著大多數物種尚未被發現和描述。(這些估計忽略了細菌,因為在定義細菌物種時存在實際問題。)在現在描述的150萬種物種中,可能有三分之二是從一個地方知道的,而許多物種是通過只檢查一個個體或有限數量的個體而知道的,因此對物種內部遺傳變異的瞭解更加有限。僅從幾個被充分研究過的物種來看,很明顯,遺傳變異可能是巨大的,而且物種之間的差異程度也不同。
山地環境特徵
山地環境與周圍低地的氣候不同,因此植被也不同。氣候的差異有兩個主要原因:海拔和地勢。海拔高度影響氣候,因為大氣溫度隨著海拔高度的增加每100公尺,下降約0.65攝氏度。山脈的起伏影響氣候,因為它們阻擋了風系統的路徑,迫使空氣在其上方上升。當空氣上升時,它變冷,導致迎風山坡上的降水量增加(地形降水量);當它下降到背風坡時,它變得更暖和,相對濕度下降,從而降低了降水的可能性,並形成了乾燥的氣候區域(雨影-降雨不易到達到的地區)。
雖然這些一般原則適用於所有山區,但特定的山區氣候各不相同。例如,沙漠地區的山區很少下雨,因為空氣幾乎總是太乾燥,在任何情況下都不允許降水,例如撒哈拉中部阿爾及利亞南部的阿哈加山脈。緯度也會影響山區氣候。在赤道地區的山區,雖然高海拔地區的溫度很低,但冬季和夏季是不存在的。在大約3500公尺以上,一年中任何一個晚上都可能形成霜凍,但在每天中午,溫度在幾乎垂直的熱帶太陽下明顯變暖,從而形成了“每晚冬天,每天春天”的當地氣候。例如,在秘魯海拔4760公尺的地方,一年中每個月的平均最低溫度約為-2°C,平均最高溫度為5至8°C。
相比之下,溫帶地區的山脈有強烈的季節性。在樹線以上的夏季,溫度高到足以讓植物生長的溫度只有大約100天,但這段時間甚至在晚上也幾乎沒有霜凍。然而,在漫長的冬季,晝夜氣溫可能保持在零度以下。在溫帶而非熱帶山區,積雪和這種降水可能引起的現象,如崩塌,是重要的生態因素。
小氣候變化在山區也很重要,由於降水量和太陽能接收量的變化,陡坡的不同方面表現出截然不同的條件。在溫帶地區,北半球朝南和南半球朝北的山坡比相反的山坡要暖和得多。這會直接或間接地影響植被;雪在地面上停留到春天的時間長度會影響植被何時出現,進而影響土地的放牧效用。即使在熱帶地區,儘管太陽處於正午垂直位置,也會出現與相位相關的氣候和植被對比。例如,在新幾內亞,朝東的山坡更溫暖、更乾燥,並且支持某些植物生長在海拔更高的地方,因為晴朗、晴朗的早晨和下午多雲的普遍模式會影響這些反差方面接收到的太陽能量。
在海拔較高的地方,山地土壤通常較淺,部分原因是土壤被冰蓋刮掉了,冰蓋形成於大約1萬年前的最後一次冰期期間,全世界大多數高山上都形成了冰蓋。土壤通常缺乏對植物重要的養分,尤其是氮。鬆散材料的快速侵蝕也很常見,並且由於凍脹、陡坡以及在溫帶地區,春季大量的融水逕流而加劇。岩石山峰和山脊上幾乎沒有土壤。然而,由於涼爽潮濕的氣候,許多山區堆積泥炭,造成當地深厚、潮濕、酸性土壤。在火山地區,火山灰也有助於土壤深度和肥力。
考慮到山脈廣闊的地理範圍和由此產生的地質和氣候變化,它們在植被方面表現出如此明顯的總體格局,這是值得注意的。除極乾旱或極寒地區外,所有地區的山地植被的主要結構特徵是樹線。(這一特徵有時被稱為林線或森林界限,儘管嚴格地說,前者是指商業規模木材樹達到的最上游,後者是指封閉的森林。)在同一座山上的斜坡之間可能會有所不同,而在低緯度,氣候變得過於惡劣,不允許樹木生長;超過這一水準,高山植被生長,主要是草本植物,如草和灌木,或低灌木。
一般來說,樹線出現的海拔高度由最熱月份的平均溫度約為10°C的高度決定,前提是濕度不是限制因素。然而,並非所有情況下都是如此;例如,在一些有一年的生長季節的熱帶地區,森林可以在比這稍冷的條件下生長。然而,在大多數地區,特別是在溫帶地區,這一價值仍然成立。它反映了一個基本的要求,即有足夠的光合作用來支持樹幹的生長。
一個相對狹窄的中間或混合植被帶-亞高山-通常存在于下面的森林和上面的高山植被之間。在溫帶山區的亞高山上,各種樹種的矮小、通常不能生育的個體能夠存活下來,儘管有風吹雪、霜凍和乾燥。這些變形的灌木大小的樹被稱為”krummholz”。
儘管森林讓位給高山植被的總體格局在所有緯度的山區都是常見的,但造成這種現象的因素並非所有地方都相同。在溫帶山區,生長季節的短暫是最重要的,因為在生長停止和冬季條件開始之前沒有足夠時間硬化的樹苗組織可能在冰凍時死亡。在這個溫帶地區,在冬季,破壞或殺死嫩枝或整棵樹的其他因素包括被風吹起的雪晶磨損嫩芽,雪堆上方的嫩枝乾燥,在那裡它們暴露在直接和雪反射的太陽輻射下,尤其是在冬季晚些時候,隨著太陽角的上升和雪地底下的嫩枝被雪菌感染。如果雪的絕緣層被吹離地面,也可能對根部造成凍傷。
在熱帶地區,這些現象是沒有經驗的。降雪並不局限於一個冬天,當它發生時,它通常融化得很快。因此,雪不會堆積成厚而連續的覆蓋層,除非海拔高於大多數植物生命的上限。例如,在委內瑞拉,樹線在4000公尺以下,即使在沒有人為干擾的地方,但實際上永久性的積雪直到大約5000公尺才出現,那裡沒有維管植物存活。熱帶地區的樹線是全年最高氣溫較低的結果。然而,地面附近的小氣候更溫暖,使得匍匐灌木能夠在遠高於最高樹木的高度生長。

山地生態系統植物群 Flora
北溫帶地區的山脈,如北美、歐洲和亞洲北部的山區,通常在其較低的山坡上有針葉樹為主的森林,讓位于上面的高山植被。這些山區典型的針葉樹有松樹、冷杉、雲杉和落葉落葉松。一些地區有落葉闊葉樹,樹下有各種較小的植物,特別是在潮濕的地方。例如,日本北半部海拔1400至1500公尺的山脈有亞北極針葉林帶,主要樹木均為冷杉屬、雲杉屬和落葉松屬的針葉樹。石南花、罌粟和大蘿蔔屬植物是生長在這些森林中的一些其他植物。在一些地區發現了沼澤地植被,以苔蘚泥炭蘚為主。樺樹(樺木)在森林的上限處環繞著森林,佔據著有燃燒歷史的區域。在北美洲西北部的太平洋上,松樹、雲杉和冷杉通常是主要的喬木林。白楊(Populus tremuloides)分佈在一些地方,特別是那些有干擾歷史的地區。榿木(Alnus)見於雪崩痕跡,柳樹(Salix)是潮濕地區的重要物種。羽扇豆(Lupinus)、毛茛科白頭翁屬(pasqueflowers,銀蓮花)和石南科的各種雛菊和低矮灌木都是生長在樹下和樹線附近草地上的豐富植物區系的例子。
南溫帶山區的樹系森林在任何地點都只由一種或極少數不同類型的樹所支配;所涉及的樹木通常是闊葉樹,而不是針葉樹。例如,大多數澳大利亞山區都有以桉樹為主的林分,儘管長期的廣泛燃燒歷史可能在某種程度上導致了這種耐火樹的突出。在紐西蘭、阿根廷和智利,樹系通常不包括任何種類的樹。
相比之下,在熱帶地區,物種多樣的森林可以被描述為發育不良的常綠雨林,通常生長到樹木生長的最高極限。新幾內亞、東南亞和東非就是這種情況;然而,在熱帶安第斯山脈的部分地區,單種(薔薇科)的多花薔薇通常生長在高於所有其他樹木的高度,尤其是在石楠(堆積在懸崖底部的岩屑)上。
在樹線以上,高山植被包括各種不同的亞型,包括草原、沼澤地、低荒原和佔據縫隙的植被。例如,在日本中部海拔2500公尺以上的高山上發現了無樹的高山植被,北海道北部的海拔下降到1400公尺。日本石松、石南和禾本科植物尤為突出。像這種高山植被中的大多數其他植物一樣,這些植物在其他山地、北溫帶地區的高山地區有近親。石松的匍匐灌木在山脊上形成一公尺高的稠密矮灌叢;在潮濕的地方,它們與榿木和花楸屬植物的落葉灌木混合。其他高山群落佔據潮濕的地方,在那裡高高的草甸或沼澤地通常擁有大量的小報春花屬植物(報春花)。矮小的矮小灌木,尤指石南科的成員及其近親熱帶山地植被概況。
相反地,在積雪堆積的地方,積雪在有遮蔽的地方堆積得很深,一直持續到春末或夏末,積雪群落以石楠科植物或莎草(苔草屬植物)為主,還有許多其他小型植物。阿爾卑斯沙漠在日本的高山地區也很普遍,這些地方的土壤因最近的火山活動而變得不穩定。雖然在這些地方生存的植物是多種多樣的,但一些,如紫羅蘭,是這些惡劣的棲息地的典型。
值得注意的是,高山植被亞型的多樣性陣列中的植物區系,例如在上面的例子中,在許多溫帶和熱帶地區,通常由大約200種不同的植物組成。此外,儘管存在廣泛的生態和地理差異,但在大多數高山地區發現了許多相同類型的植物。它們通常是由不同的,雖然相關的物種在每個地區和每一個山區在區域內。龍膽、車前草、毛茛、石南、禾草和莎草都是很常見的例子。
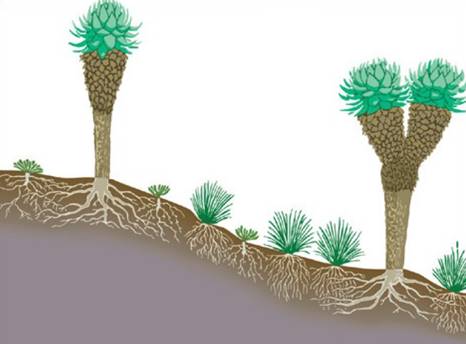
然而,高山植物區系和植被結構都具有一定的區域性。一個突出的例子是在幾個熱帶高山上發現的大莖蓮座植物,但不是溫帶的。這些是高達3公尺(約10英尺)或更高的巨型草本植物;它們有持久的枯葉基部,能使莖的含水組織免受幾乎每晚在其非常高(高達4300公尺[約14100英尺])的乾燥環境中發生的冰凍條件的影響。在安第斯山脈北部(Espeletia和Puya)和東非山區(Dendrosenecio和Lobelia)也發現了類似但不相關的莖叢植物,夏威夷、爪哇和喜馬拉雅山也有類似的例子。這種在不同大陸上處於相同環境壓力下的不同物種之間出現相同的特徵是趨同進化的一個例子。 冬雪從暴露的表面被吹走時,會出現暗斑病、越桔病、地溫病。
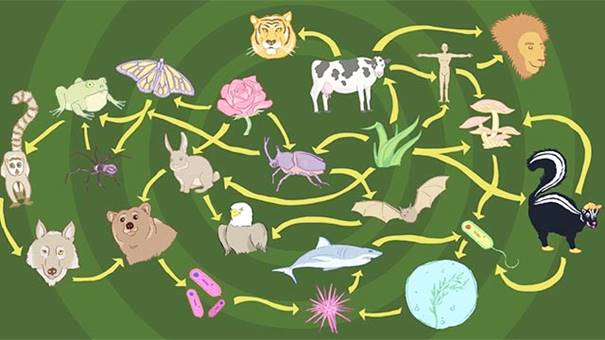
山地生態系統的動物群 Fauna
山地動物群不像同一地區的植物群那麼獨特,通常反映的是區域動物群。例如,北美山區的大型哺乳動物包括鹿、熊、狼和幾隻大型貓科動物,它們都棲息在山外的周圍地區,或在人類入侵之前就居住過。有些鳥類與山地棲息地聯繫在一起,比如加利福尼亞和安第斯山脈的禿鷲。在某些山區,蚱蜢等不會飛的昆蟲是一個有趣的特徵,這種現象在乞力馬札羅等東非山峰上尤為明顯。
由於其地形和氣候環境的多樣性,以及由於冷適應生物群的進化往往是在同一地區的不同山脈上獨立進行的,因此山區常常被認為是高度生物多樣性的中心。亞洲的高加索山脈就是一個著名的例子,而在熱帶地區,新幾內亞的山脈對生物的多樣性作出了巨大貢獻,包括約20000種植物,占世界植物區系的10%。
山地生態系統中種群和群落的發展與結構
溫帶山區的人口和群落過程,與相當相似的北極環境一樣,受到高度季節性氣候的影響。隨著冬季積雪的融化,植物會經歷生長和開花的高潮,尤其是在高山地區,整個生長季節大約在三個月內完成。地下器官中大量的食物儲備被用來快速地產生成熟、可育的嫩枝,在某些情況下,在融雪完成之前就開始生長。因為幾乎整個高山植物群都在同一時期開花,只有幾個星期,而且高山植物往往有相對較大的花,所以夏季在溫帶山區看到的花卉展示通常是壯觀的。在熱帶山區,沒有這樣一個壯觀的發展時期,與一個較長的強制休眠季節交替出現,植物全年都在生長,除非它們的發育因旱季的到來而停止。
動物活動在不同地區也有相似的季節性變化。在溫帶山區,大多數鳥類和大型哺乳動物遷徙到低海拔地區的時間很長。一些僅存的哺乳動物,如北美山區的地鼠,利用積雪下的絕緣環境,在那裡它們挖洞,以地下植物器官為食。
在熱帶山區,季節變化不太明顯,這反映在動物繁殖上。例如,新幾內亞高山上的鳥類可能全年繁殖。然而,由於沒有季節性的植物和昆蟲的生長,創造了暫時充裕的食物來源,它們產卵很少。與許多溫帶山鳥在短暫的繁殖季節通常會產下5到8個蛋相比,那裡通常只有一到兩個。
山地生態系統的生產力
山地作為植物生長的壓力大的棲息地,並不是很高產的環境。然而,高溫帶山區高山植被的生物量(一個地區的有機物幹重)可能比最初出現的要大,因為在地下以根、根莖、塊莖和球莖的形式存在的可見地上生物量超過10倍。相比之下,熱帶高山植物群的植物不需要將食物儲存在地下,而且不到總生物量的一半位於地下。
因此,山區的農業開發不是很有生產力,一般也不集約,主要局限于牛、羊和羊的光或季節性放牧。在中等強度放牧的地方,放牧對高山植被的破壞性非常大,而高山植被在環境緊張的狀態下很難應對干擾。同樣,人類在高山上的其他用途,如滑雪和其他形式的娛樂活動,會造成永久性的損害。另一個擔憂是,大氣污染物往往集中在降雪中。在溫帶地區,污染物質隨著每年的融雪進入高山系統,在這種低營養環境中可能帶來有害後果。
n 極地生態系統
俄羅斯術語“苔原”來源於芬蘭語單詞tunturi,意思是無樹的高緯度。苔原現在被用於一般意義上描述任何沒有樹木的植被的寒冷氣候景觀,包括山區(高山凍原)和北極、亞北極和南極地區。在更嚴格的意義上,苔原是指一種特殊類型的植被組合。除了岩石露頭、乾燥的山脊頂部和河流礫石壩外,大多數地區都有充分的植被,主要是矮生灌木、地衣和苔蘚。極地地區的苔原地帶與植被稀少的極地貧瘠地帶截然不同。

極地動植物的起源
與其他生物群落相比,北極苔原生物群落相對年輕,起源于更新世(距今260萬至11700年)。然而,凍土帶的個別動植物物種可能最早出現在中新世晚期(1380萬至530萬年前)或上新世早期(530萬至360萬年前)。在上新世中期(360萬年前),埃爾斯公尺爾島和格陵蘭島北部是最北端的陸地地區,那裡有針葉林。大多數古生態學家認為,苔原植物群是在中新世(2300萬至530萬年前)大陸漂移到高緯度和低緯度地區時,從針葉林和高山地區的植物演化而來的。
相比之下,南極洲大陸自古近紀早期以來,即大約6000萬至4000萬年前,一直被廣闊的海洋與其他大陸板塊隔離開來。在它分離之前,它和澳大利亞、南美洲、印度半島和非洲一起作為岡瓦納大陸的一部分而存在。這種長期的分離阻礙了南極陸生動植物的建立和發展。阻礙陸地生物進化的其他重要因素是惡劣的氣候、更新世冰川時期完全吞噬了整個大陸的冰蓋,以及目前數量有限的無冰陸地區域,主要局限於海岸邊緣和努納塔克(被冰蓋包圍的山峰)。因此,南極陸生動植物很少。南極洲半島延伸至南緯63度,幾乎是南極所有植物生長的地方。
然而,南極不僅包括大陸本身,還包括位於南極輻合帶內的島嶼,那裡向北流動的冷地表水與溫暖的亞北極水域相遇。大多數南極島嶼,由於其位置在季節性浮冰之外,受到的海洋影響要比可比的北極島嶼強得多。這些島嶼的動植物發展很差,主要是因為它們與潛在的陸地生物源隔絕。南喬治亞島是這些島嶼中最大的一個,位於火地島以東2000公里,南緯54度至55度,面積3756平方公里。在高海拔地區和廣闊的冰川地區,強降水和高山是常年積雪的原因。
然而,在有可能形成土壤的海平面附近,形成了獨特的南極苔原植被,主要是草叢形成的草和苔蘚。目前只有26種維管植物,而且沒有本土哺乳動物,儘管19世紀初挪威捕鯨者引進的馴鹿已經建立了野生種群。馴鹿的放牧通過減少或局部消除最喜歡的物種而改變了植物群落。
極地環境特徵
極地環境的特點氣候、基底、海拔高度、坡度、暴露以及與其他大陸的接近性決定了極地地區存在的動植物的複雜多樣性。正是這些因素在夏季的相互作用決定了溫度是否足夠溫暖,濕度是否足以讓植物生長。根據生態標準,高北極區與低北極區有所區別,包括生長季節較短,夏季涼爽,動植物種類明顯減少。
北半球苔原帶的南界可能從加拿大哈德遜灣南端的55°N沿著阿拉斯加的白令海海岸和俄羅斯遠東延伸到加拿大麥肯錫河下游、西伯利亞中部的哈坦加河和斯堪的納維亞北部的70°N以上。這一限制通常與每年75至80千焦/平方cm的太陽淨輻射等值線一致,這與7月10°C的等溫線非常接近。然而,這一邊界的局部變化發生在北美和歐亞大陸,那裡的山脈或暖流的影響使森林向北滲透到輻射量只有每平方cm67千焦耳的地區。
在典型的苔原和極地貧瘠地區,夏季的平均氣溫可能只有2到5℃,植物生長季節通常不到100天。雖然高緯度地區的夏季短暫,但夏季的白天卻很長。在北極圈以上的緯度地區(北緯66°33',至少有一部分夏天,太陽每天24小時都在地平線以上。這種持續的陽光使植物能夠優化光合作用,而無需通常的夜間呼吸成本。
凍土帶和極地貧瘠地區的年總降水量很低,一般在每年100至1000mm之間。在沿海和高海拔地區,降水量通常最大。例如,在格陵蘭島和南極洲以及加拿大北極群島的高海拔地區,形成了永久性的冰蓋。苔原地區的低降水特徵與低緯度地區的沙漠相當。然而,這些地區不同於真正的沙漠,因為冬季和夏季的低溫限制了蒸發。因此,融雪和夏雨帶來的有限水分中,大部分可用於植物生長。此外,大多數凍土帶和極地貧瘠地帶下麵是永久凍結的土壤(永凍層),除了每年夏季融化的薄表層(活躍層)。永凍層限制了排水,並保留了活躍層內植物生長所需的水分。這會導致濕地的形成,就像北極沿海平原的水準區域一樣,那裡有大量的濕地,是水生植物、無脊椎動物、水禽和濱鳥的家園。
地球周圍的氣團將工業污染物從溫帶地區輸送到北極和南極。污染物正逐漸集中在這些地區,人們對高緯度地區生命可能造成的後果越來越關注。由於工業向大氣中排放氟氯烴而導致極地地區臭氧層變薄,可能對動植物的生命造成長期的影響。以前被臭氧層截獲的紫外線輻射對大多數生命都是有害的,因為它抑制光合作用,增加DNA突變率,並損害動植物的表皮組織。
北極動植物群 B iota
在樹木的北部邊界存在一個過渡帶,針葉林與無樹苔原植被交錯。在北美,白雲杉和黑雲杉與凍土帶交界,而在西伯利亞和北歐,落葉松是主要的樹種。棉白楊(胡楊種)通常沿著主要河流滲透到低北極的苔原景觀中。低北極的主要植被類型包括低灌木苔原,主要是柳樹(Salix)和矮樺樹(Betula);高灌木苔原,主要是柳樹、樺樹和榿木(Alnus);以及莎草和矮灌木的組合,如拉布拉多茶(Ledum)、藍莓和蔓越莓(Vaccinium)等,在潮濕的地方有烏鴉莓和北極石南。墊狀植物(旱獺和虎耳草屬)在風刮的高地上很常見。地衣和苔蘚是某些地區地表覆蓋物的重要組成部分。在低北極地區,除了岩石露頭、乾燥的山脊頂部、河流礫石壩和碎石坡(在休憩角有岩石碎屑堆積的斜坡)外,大多數陸地表面都有植被。
高北極地區的植被不如低北極地區的豐富,僅包含低北極地區發現的一半左右的維管植物物種。例如,在北美洲的低北極地區發現了600多種植物,但在埃爾斯公尺爾島北部和83°N以北的格陵蘭島的極端高北極地區,生長的維管植物不到100種。較短的生長季節,涼爽的夏季,乾燥的條件,以及這些大陸與大陸植物群的距離,解釋了這種差異。北極地區40%以上的維管植物是環極分佈的。苔蘚在高北極植物群落中的重要性增加,灌木種類明顯減少,只有少數匍匐的柳樹、矮樺樹和其他矮生灌木保留下來。然而,匍匐的柳樹仍然是植物群落的重要組成部分,即使在最北部的陸地地區也能保持一定的冬季積雪。莎草苔蘚草甸出現在由融雪澆水的穀底有限的潮濕地點。高地比較乾燥,地表覆蓋物比較稀疏,在海拔較高或水分不足的地方,地表覆蓋物會併入極地沙漠。草地,偶爾匍匐的柳樹,和墊狀的旱地出現在高地上,是極地貧瘠地區的主要植被。
真正的極地沙漠一般出現在北冰洋沿岸的沿海地區和北極海拔幾百公尺的地區,那裡的土壤尚未發育,無霜期和土壤濕度不足以滿足大多數植物的生長。偶爾生長在那裡的植物通常會生長在霜凍裂縫中,這些裂縫捕獲了吹雪和更細的風吹土壤材料。適應這些條件的植物包括北極罌粟、一些燈心草)、小薩克斯風草(虎耳草)和其他一些形成蓮座叢的草本物種。北極罌粟和其他一些適應高北極地區的開花草本植物的花是具有太陽光效應的(在太陽的作用下轉動)。它們抛物線形的花朵跟蹤太陽的日常運動,從而將太陽熱量集中在發育中的卵巢上,使降落在那裡的授粉昆蟲變暖,並加速胚胎種子的生長。
北極生態系統缺乏溫帶和熱帶生態系統特有的物種多樣性和豐富性。在兩極地區,動物和植物物種的數量隨著緯度的增加而減少。北極凍原和極地貧瘠地帶的脊椎動物物種僅限於哺乳動物和鳥類;那裡沒有兩棲動物或爬行動物。北極地區有大約20種哺乳動物和100多種鳥類。大多數是環極分佈的單種或近緣種,例如北美馴鹿和歐亞大陸的家養和野生馴鹿屬於同一物種,Rangifer tarandus,而歐亞北極地區的旅鼠與北美洲北部和格陵蘭島的旅鼠關係密切,但又截然不同。北極哺乳動物區系的這種相似性是更新世冰川海平面較低的結果,當時廣泛的陸地連接,即白令陸橋,連接了今天的阿拉斯加和西伯利亞。
一些北極哺乳動物群主要是食草動物,如馴鹿和馴鹿、麝牛(Ovibos moschatus)和北極狐(Alopex lagopus),以及北極野兔(Lepus)和有領和棕色旅鼠(Dicrostonyx和Lemmus)的物種很少出現在北極以外,並且適應於這種環境中的生活。其他動物群,如地松鼠(Spromophilus)、田鼠(田鼠)、鼩(Soricidae)和紅狐(Vulpes),以及貂(Mustela erminea)、狼獾(Gulo Gulo)、狼(Canis lupus)和棕熊(Ursus arctos)是其他生態系統常見的動物,但在整個北極地區廣泛分佈。其他一些典型的溫帶物種已經向北滲透到低北極地區,那裡有合適的棲息地。北美的駝鹿(Alces Alces)和雪鞋兔(Lepus americanus)就是例子,它們向低北極移動可能是由於
氣候變暖,柳樹和其他灌木的增加,尤指在河岸棲息地。在那裡,北方森林地區的山脈一直延伸到北極,就像在北美西北部一樣,西伯利亞的山地綿羊(Ovis)和土撥鼠(旱獺)的物種已經擴展到了北極地區。
在北緯80°以上的極端高北極地區,僅包括加拿大北極地區的阿克塞爾-海伯格和埃爾斯公尺爾群島、格陵蘭島最北端、斯瓦爾巴特島北部和弗朗茨-約瑟夫島的部分地區,只有少數哺乳動物能夠維持生存的種群。在加拿大的高北極地區,麝牛、梨鹿、北極兔和有領旅鼠是唯一的哺乳動物食草動物,它們的捕食者狼、北極狐、狼獾和貂也存在。在格陵蘭島北部也發現了同樣的物種,歷史上馴鹿和狼獾可能都不存在。只有北美馴鹿和北極狐原產於斯瓦爾巴群島,只有北極狐出現在弗朗茲·約瑟夫的土地上。在所有這些高北極地區,北極熊(Ursus maritimus),一種主要捕食海豹的海冰生物,偶爾會在陸地上被發現,在那裡雌性動物棲息繁殖幼崽,或在那裡吃草(很少)或捕食陸地哺乳動物或築巢鳥類。
北極陸生鳥類區系僅包括少數幾種常住物種,其中包括斑尾雉(Lagopus species)、雪鴞(Nyctea scandiaca)、gyrfalcon(Falco rusticolus)和烏鴉(Corvus corax);其餘物種僅在夏季出現在北極,用於繁殖和繁殖幼鳥,遷徙至溫帶、熱帶或海洋地區冬季南部緯度更高。雖然鳥類的飛行能力使它們能夠在北極圈內佔據哺乳動物基本上無法進入的與世隔絕的棲息地,但它們在整個北極的分佈與它們越冬地區的位置和每年的遷徙路線密切相關。這些遷徙路線,特別是那些濱鳥和水禽的遷徙路線,通常沿著大陸的海岸線,然而有些物種跨越廣闊的水域。然而,北大西洋對物種的環極混合提供了部分障礙,因此,北美洲西部和歐亞大陸東部北極地區的鳥類區系之間的相似性比歐洲、北美東部和格陵蘭島北極地區的物種更為相似。
濱鳥、水禽和雀形科雀形目鳥類(雀、雀、雀)是北極地區築巢最豐富的物種。潮濕的莎草草甸通常與湖邊、河口和海岸相連,是濱鳥和水禽的最佳築巢棲息地。雀形目鳥類的築巢密度在凍土帶南緣的灌木群落和河岸棲息地中最高;在高北極地區,它們迅速下降。在這一類群中,只有紅豆(acanis)和雪旗(plectrophenaxnivalis)將它們的範圍擴展到最北部的陸地地區。
除了常住物種外,通常在北極築巢的猛禽還包括遊隼(Falco peregrinus)、粗腿鷹(Buteo lagopus)、短耳貓頭鷹(Asio flammeus),以及在山區,北美的金鷹(Aguila Chrysetos)和格陵蘭和格陵蘭島的白尾鷹(Italiacetus albicilla)歐亞大陸。jaegers(Stercorarius物種)一年中大部分時間在海上度過,它們在凍土帶和極地貧瘠地帶築巢,在繁殖季節捕食旅鼠、蛋和其他鳥類的雛鳥。原螯蝦科(fulmars)、落葉松科(海鷗和燕鷗)和海雀科(海雀、海雀、多夫鳥和海雀)的海鳥,依賴于海洋食物,通常在北極的海岸懸崖上殖民地築巢。這些築巢群通常是在靠近上升流的海面上發現的,在那裡鳥類以無脊椎動物和魚類為食。在高緯度的北極地區,上升的洋流會在被稱為波利尼婭的浮冰中形成開闊水域;這使得海鳥能夠在北緯75°以上的地方覓食和築巢。




南極動植物群 B iota
南極洲的植物區系主要由土壤和淡水藻類、地衣、苔蘚、真菌和僅有的兩種本地維管植物組成。陸生動物群包括一些無脊椎動物種類:原生動物、輪蟲、線蟲、緩步動物、彈尾蟲(原始無翅昆蟲)和一種蟎類。這些生命形式主要局限於潮濕的苔蘚層。沿海地區海洋哺乳動物和鳥類的多樣性以及相關的浮冰依賴於鄰近海域的海洋食物鏈。
在南極洲,獨特的內石器(石頭居住)生命形式(藍藻)出現在多孔岩石表面內部和下方。這些藍藻可以在維多利亞州南部乾旱的山谷中發現,在那裡它們適應了長時間的休眠,直到罕見的情況下融化的雪提供了生命過程所需的水分。雖然這是一個極端寒冷的沙漠環境,但在炎熱沙漠的岩石中也有類似的生命形式。
極地生態系統中種群和群落的發展與結構
極地地區動植物物種多樣性低,導致北極和南極生態系統缺乏複雜性。植物可以生長,昆蟲和其他無脊椎動物也可以活躍,這一短暫的夏季導致了這些生態系統的生產力低下和相對簡單。此外,較冷的溫度限制了植物生長所必需的土壤養分通過分解有機物質、分解母岩和土壤微生物固氮釋放的速率。
生態系統簡單化的後果是缺乏穩定性。動物在凍土帶和極地貧瘠地帶往往會經歷大範圍的種群波動。這些波動是由週期性的極端天氣和草食植物和捕食者-獵物關係的不平衡引起的。與高度複雜的溫帶和熱帶生態系統中的物種相比,每種物種在北極生態系統的營養動態中發揮著更為主導的作用。旅鼠在其種群週期的高峰期(每公頃高達200只),繁殖成功率高,捕食者數量迅速增加,如北極狐、貂、雪鴞和其他物種。相反,當旅鼠數量較低(每公頃不到1只)時,捕食者之間的繁殖停止,捕食者數量急劇下降。
在高緯度的北極地區,動物種群生活在接近其生物學極限的地方。梨馴鹿或麝香牛可能會受到極端天氣的影響而在當地滅絕。在有利的條件允許鄰近種群建立之前,物種的重建可能不會發生,這可能需要幾十年的時間。事實上,在格陵蘭島最北端,儘管馴鹿在近代歷史上從未出現過,但鹿角和骨頭表明它們至少在7000年前定期出現。
雪對凍土帶和極地貧瘠地區動植物群落的特徵和分佈起著重要的作用。冬天很長,落下的有限的雪通常在整個季節都不會融化。然而,積雪並不穩定。這些無樹景觀特徵的強風重新分佈了積雪,將其從景觀凸面上移除,並將其堆積成凹坑、灌木斑塊或山脊頂部和巨礫背風方向的漂移物。風也會壓縮雪,增加雪的密度,從而使動物在雪表面更容易移動,同時也會阻礙它們在下面挖掘食物的能力。因此,雪會影響大型食草動物的位置、食物選擇和能量消耗,它們必須穿過冬季的景觀來覓食。積雪較深的地區對小型齧齒動物很有吸引力,它們在積雪的絕緣層下的地面上覓食和築巢。
凍土帶和極地貧瘠地帶的積雪分佈模式也是植被分佈的主要決定因素。大多數植物需要積雪來保護它們免受冬季極端寒冷和乾燥的影響。積雪被吹走的地區是第一個在夏季開始植物生長的地區,而那些積雪融化緩慢的地區只支援那些適應較短無雪季節的植物。然而,冬季積雪通常是植物夏季生長的主要水分來源。因此,有風的暴露和很少的積雪通常遭受夏季乾旱的限制植物生長,而雪床植物群落在整個夏季都得到了良好的澆灌。
極地生態系統的生產力
在低北極地區,植被覆蓋了80%到100%的陸地面積。維管植物在短暫的北極夏季的連續日光下快速生長,是這些植物群落在如此高海拔地區相對較高生產力的基礎。與森林生物群落相比,苔原植物的所有地上生產力都接近地面水平,因此脊椎動物很容易獲得這種生產力。因此,大量北美馴鹿和鵝在夏季在低北極地區局部放牧,但在飼料品質下降且其可用性受到風雪覆蓋的限制時從北極遷徙到冬季。儘管苔原系統在短暫的北極夏季經歷了植物和動物生產力的激增,但在漫長的冬季,只有少數動物物種在北極保持活躍。食草動物依賴前一個夏天的植物生產,而食肉動物則依賴於他們的食草動物獵物。凍土帶和極地貧瘠地區的年總生產力比大多數溫帶或熱帶生態系統低幾個數量級。
脊椎動物的草食可能導致苔原植被的變化,通過選擇性的取食,踐踏和通過排泄循環利用土壤養分。在馴鹿、麝牛、旅鼠和鵝集中它們的放牧活動的地方,它們實際上可以增加苔原植被的產量。這是由於大量的一年生植物生長被移除,植物新生長的潛在部分暴露在陽光下,土壤受到越來越高的熱量,以及通過消化和排泄加速有機物質的循環,從而將養分釋放到土壤中。
