
河流廊道復育 www.epa.url.tw
Stream Corridor Restoration Handbook 林雨莊編譯
United States Department of Agriculture
The Federal Interagency Stream Restoration Working Group
第一章 河流廊道概況
第二章:廊道過程、特徵和功能
第三章:影響河流廊道的干擾
第四章:復育工作組織和問題識別
第五章:制定目標和恢復方案
第六章:實施、監控、評估和適應
第七章:廊道條件分析
第八章:河流修復設計
第九章:恢復實施、監測和管理
第二章:廊道過程、特徵和功能
2A 水文和水力過程
2B 地貌過程
2C 物理及化學特性
2D 生物群落特徵
2E 河流廊道
2F 河流功能與動態平衡
第二章 河流形成過程與特徵
2A 水文水理過程
� 河流量來自何處?
� 哪些河流程影響或涉及河流河流?
� 水的河流動速度,速度,深度,頻率和時間?
� 城市河道的水文情 況 如何不同?
2B 地貌過程
� 哪些因素會影響河道橫截面和河道輪廓?
� 水和沉積物如何相關?
� 沉積物來自何處以及如何向下游輸送?
� 什麼是均衡河道?
� 橫截面和剖面中的河道應該是什麼樣的?
� 如何進行河道調整?
� 什麼是洪氾區?
� 河流與洪氾區之間是否存在重要關係?
2C 物理化學特性�
� 水的主要化學成分是什麼?
� 物理棲息地和關鍵化學參數之間有哪些重要關係?
� 化學和物理參數如何對河流中的水生生物起關鍵作用?
� 河流水質中的天然化學過程是什麼?
� 環境干擾如何影響水質的化學特徵?
2D 生物群落特徵�
� 河道的重要生物成分是什麼?
� 在河道內可以找到哪些生物活動和生物?
� 河道的結構如何支持各種生物群?
� 有助於河道生物多樣性的水生系統的結構特徵是什麼?
� 河道內發生了哪些重要的生物過程?
� 魚在河廊復育中有什麼作用?
2E 河流功能與動態平衡
� 河流的主要生態功能是什麼?
� 隨著時間的推移,這些生態功能如何保持?
� 河流是否穩定? � 這些功能是否相關?
� 河流如何回應水流、植被、泥砂等自然力 ( 即動態平衡 ) ?
第 2A 節:水文水理過程
了解水流入和河流過河道是如何進行修復的關鍵。河流速、水量、水深、持續性都是重要的基本關鍵。
第 2B 節:地貌過程
本節將基本的水文過程與物理、地貌功能特徵相結合。水流受到河道、洪氾區和高地內土壤和沖積特徵的影響。河流攜帶的沉積物數量和種類在很大程度上決定了其平衡特徵 ( 大小、形狀和輪廓 ) 。成功的河流廊道復育,無論是主動 ( 需要直接變化 ) 還是被動 ( 管理和消除干擾因素 ) ,都取決於對水和沉積物如何演變的過程理解。
第 2C 節:物理化學特性
河道中的水質、水量通常是河流復育的主要目標,將其改善到生物所需條件,或者維持水質、水量條件。復育應考慮明顯或不明顯的物理和化學特性,河道的功能和過程至關重要。為實現復育目標必須改善土壤或水質化學條件,涉及河流管理或改變景觀中的元素。
第 2D 節:生物群落特徵
在河道中居住的魚類,野生動植物、植物和人類是復育過程中需要考慮的因素。典型的目標是復育、創造、增強或保護棲息地以使生存、繁殖條件優化。更要了解水是如何河流動的,如何運輸沉積物,以及地貌特徵和過程是如何演變的 ; 然而,成功復育的先決條件是了解系統的生命部分以及物理和化學過程如何影響河道。
第 2E 節:河流生態功能和動態平衡
河流的六大生態功能是:棲息、引導,阻隔、過濾、來源和匯集。河流生態系統的完整性取決於這些功能的運行情 況 。本節討論這些功能以及它們與動態平衡的關係。

圖 2.1 :運動中的河流廊道。過程、特點,功能塑造了河流廊道面貌。
2A 水文水理過程
水文循環描述的水從降雨轉移到地表水和地下水,以存儲和逕流的連續體,並通過蒸騰和蒸發 ( 圖 2.2) 最終返回到大氣中。
降雨使水返回地表。雖然大多數水文過程都是根據降雨事件來描述的,但融雪也是重要的水源,特別是對於起源於高山地區和大陸地區的河流而言。
降水的類型通常是濕度和氣溫的一個因素。地形起伏和相對大型水體的地理位置也會影響降水的頻率和類型。在溫度適中、地勢低平的沿海和低緯度地區,豪雨的發生頻率更高。在高海拔地區和中緯度地區,氣溫較低時,降雪頻率更高。
一旦到達大地,降雨有三個去向,它可以蒸發回歸大氣 ; 下滲進入土壤 ; 或者從地表河流入河流湖泊、濕地或其他水體。這三條路徑都在決定水流入、河流過和河流入河道的過程中發揮作用。
本節分為兩個小節。第一小節側重於橫向尺寸的水文和水理過程,即水從陸地進入河道的運動。第二部分集中于水在縱向上的河流動,特別是在河道中的水流。

圖 2.2: 水文循環。包含降水、地表水和地下水、儲存和逕流 , 並最終回到大氣中是一個持續的循環圈。
2A-1 河流整體的水文水理過程
水文循環的關鍵點在本小節中作為組織標題:
■ 攔截,蒸騰和蒸發。
■ 滲透,土壤濕度和地下水。
■ 逕流。
2A-1.1 截留,蒸騰與蒸發
在上空的降雨中約有三分之二以上蒸發到大氣中,而不是作為河流入海洋的水流排出。水文循環的 � 捷徑 � 是由於兩個過程,攔截和蒸騰而發生的。
攔截、截留
一部分降雨從未到達地面,因為它被植被和其他自然和構造的表面攔截。以這種方式截獲的水量由地上表面上可用的攔截存儲量決定。在植被區域,儲存是植物類型和葉,枝和莖的形態和密度的功能 ( 表 2.1) 。影響森林地區儲存的因素包括:
■ 葉形。針葉樹針比葉子更有效地保持水分。在葉子表面上,液滴一起運行並滾落。然而,針頭保持液滴分離。
■ 葉紋理。粗糙的葉子比光滑的葉子儲存更多的水。
■ 一年中的時間。無葉期對冠層的攔截潛力小於生長期 ; 但是,在此期間,葉凋落物會產生更多的存儲地點。
■ 垂直和水平密度。降雨必須穿透的植被層越多,到達土壤的可能性就越小。
■ 植物群落的年齡。隨著年齡的增長,一些營養林變得更加密集 ; 其他人變得不那麼密集降雨的強度,持續時間和頻率也會影響攔截量。
一場大雨的開始首先填滿了樹冠枝葉上的攔截儲存區。隨著大雨的持續,樹冠枝葉的儲存區容納不下的水被轉移了,散置的水滴到樹下灌木和草地,並填滿那裡的儲 存區。過程再重複後,雨水落到達最底的落葉屑層。從落葉層排出的水或滲入土壤,或隨著地表逕流向地形低窪處移動。
先前的大雨若不再持續,蒸發開始移去攔截儲存的水分,包含樹葉上的、草地上的、淺層土壤間隙的。蒸發率因環境、氣溫、地區而異,每個地方都不同,台灣地區的蒸發 + 蒸散率約為 29% ,土壤入滲量約為 18% ,地表逕流量 71% 。 ( 楊萬全, 2000)
在植被很少或沒有植被的地區,攔截通常是微不足道的。裸露的土壤或岩石有一些小的不可滲透的凹陷,它們起著相互作用。
在大城市地區攔截是非常重要的。雖然城市排水系統的設計是為了快速將雨水從不透水的表面排出,但都市景觀中有豐富的儲存場地。這些包括平坦的屋頂、停車場、坑洞、裂縫和其他粗糙的表面,這些表面可以截留並容納水以進行最後蒸發。
蒸騰與蒸發
蒸發是水體受熱,水分子進入大氣中,水庫、埤塘、河流、水稻田都會直接蒸發,植物枝葉表面、土壤孔隙地水分子進入大氣的也是蒸發。蒸騰與蒸發不同,植物根系從土壤吸收了水分,從葉片上的氣孔擴散到大氣中才是蒸騰。
雖然土壤中的水分也會蒸發。但是由於毛細管作用使土壤中的水分蒸發很緩慢,大部分土壤中的水分子會涵蓄、握持、緩慢下滲或橫向滲河流往河道。
只要暴露在大氣中,水就會蒸發。基本上這個過程包括:
■ 蒸發水分子的運動在大氣中產生蒸汽壓,蒸發速率與水體表面積及蒸汽壓差成正比。當蒸氣壓均衡時,就不會再發生蒸發,直到別處的空氣對河流進來水體上空。因此蒸發率根據緯度、季節、日照時間、雲量、濕度、大氣壓和風能而變化 部分降雨永遠不會到達地面 , 因為它被植被和其房屋等截獲,直接被蒸發。
■ 蒸發,這兩個過程通常結合並標記的蒸散。蒸發蒸騰可以控制水平衡,可以控制土壤水分含量、地下水補給和河流量。

圖 2.3: 森林降雨的典型途徑。
圖 2.4: 台灣降雨量與蒸發量。中南部降雨集中於 7 至 10 月,冬春兩季乾旱。


圖 2.5: 土壤剖面。水通過重力和毛細管作用被吸引到土壤中的毛孔中。
表 2-1 :各種植被類型截留降雨量的百分比

2A-1.2 滲透,土壤浸潤與地下水
未攔截或地表逕流河流入降雨移動到土壤中。在那裡,它可以存儲在上層或向下移動通過土壤剖面,直到它到達被稱為潛水區的水完全飽和的區域。滲透對土壤表面的仔細檢查顯示,數百萬個沙子,淤泥和粘土顆粒被不同大小的河道分開 ( 圖 2.5) 。這些大孔包括裂縫,腐爛的根和蟲洞留下的 � 管道 � ,以及土塊和土壤顆粒之間的孔隙。通過重力和毛細管作用將水吸入孔隙中。重力是水流入最大開口的主要力量,例如蠕蟲或根孔。毛細管作用是水進入具有非常細孔的土壤的主導力。
浸潤滲透
這些孔隙的大小和密度決定了水進入土壤的速度。孔隙度是用於描述土壤顆粒之間的空間佔土壤總量的百分比的術語。當所有這些空間都充滿水時,據說土壤已經飽和。土壤特性如質地和傾斜度 ( 鬆散度 ) 是 決定孔隙度的關鍵因素。粗糙的沙質土壤和土壤由鬆散的聚集體通過有機物質或少量粘土固定在一起,具有大的孔隙,因此具有高孔隙率。緊密堆積或粘土的土壤孔 隙率低。滲透是用於描述水進入土壤孔隙的術語。滲透率是水,超過給定的時間長度滲入土壤的量。水滲入土壤的最大速率被稱為土壤的滲透能力。如果降雨強度小 於滲透能力,水以等於降雨率的速率滲入土壤。如果降雨率超過滲透能力,多餘的水要么被留在土壤表面的小凹陷中,要么作為地表逕流向下行進 ( 圖 2.6) 。以下因素對於確定土壤的入滲率非常重要:
■ 易於通過土壤表面進入。
■ 土壤中的儲存能力。
■ 通過土壤的傳播率。
具有天然植被覆蓋和落葉的區域通常具有高滲透率。這些特徵保護表面土壤 poi4 空 間不被雨滴飛濺產生的細小土壤顆粒堵塞。它們還為蠕蟲和其他穴居生物提供棲息地,並提供有機物質,幫助將細土顆粒結合在一起。這兩種方法都增加了孔隙率和 滲透速率。在整個暴雨期間,滲透率不是恆定的。率是通常在暴雨開始時高,但作為重力供給存儲容量被填充迅速下降。通常在暴雨中 1 或 2 小時達到較慢但穩定的滲透速率。有幾個因素在 volved 在此穩定化過程,包括以下內容:
■ 雨滴破壞土壤聚集體並產生更細的物質,然後阻擋表面上的孔隙並減少進入的容易程度。
■ 水填充細孔空間,降低存儲容量。
■ 潤濕的粘土顆粒膨脹並有效地減小了孔隙空間的直徑,從而降低了傳輸速率。
暴雨過後,土壤逐漸排水或乾燥。
但是,如果在乾燥過程完成之前發生另一場暴雨,新水的存儲空間就會減少。因此,在分析可用存儲時,先前的潮濕條件很重要。

圖 2.7: 各種土壤的保持水分特性。保持水的特性因紋理而異。對於細沙地 , 孔隙度、 0.45 和場容量 0.20 之間的近似差異為 0.25, 這意味著未填充的孔隙空間是土壤體積的 0.25 倍。場容量和枯萎點之間的差異是未填充孔隙空間的度量。 Source: Dunne and Leopold 1978.
土壤濕度
暴風雨過後,水會因重力而從上層土壤中排出。然而,土壤保持濕潤,因為一些水通過表面張力保持緊密地保持在細孔和顆粒周圍。這種情況稱為田間容量,隨土壤 質地而變化。與孔隙率一樣,它表示為體積比例。孔隙度和田間容量之間的差異是未填充孔隙空間的量度 ( 圖 2.7) 。 然而,現場容量是一個近似數字,因為在潮濕的土壤中以低速繼續引力引河流。在蒸發蒸騰的背景下,土壤水分是最重要的。陸生植物依賴於土壤中儲存的水。由於 它們的根從逐漸變細的孔隙中提取水分,土壤中的水分含量可能會低於田間容量。如果土壤水分沒有得到補充,根最終會達到這樣的程度,即它們無法產生足夠的吸 力來提取緊密保持的間質孔隙水。在這一點上,根據土壤特性,其改變土壤的含水量,被稱為永久萎蔫點,因為植物不能再從高到足以跟上蒸騰的需求率土取水,造成植物枯萎深層滲透是指通過農作物根區以下的水量,減去從根區以下向上河流動的水量 (Jensenetal.1990) 。
地下水
孔隙開口的大小和數量也決定了土壤剖面中水的運動。重力原因
水垂直向下移動。這種運動很容易通過較大的毛孔。由於粘土顆粒膨脹或孔隙填充導致孔隙尺寸減小,因此河流動阻力更大。毛細管力最終接管並導致水向任何方向移動。
水將繼續向下移動,直至達到完全飽和的水,潛水區或飽和區 ( 圖 2.8) 。潛水區的頂部定義了地下水位或潛水面。在地下水位上方是一個稱為毛細管邊緣的區域,因為這個區域的毛孔充滿了毛細管的水而得名。
在具有微小孔隙的土壤中,例如粘土或淤泥,毛細管力很強。因此,毛細管邊緣可以從地下水位向上延伸大的距離。在具有大孔 的砂岩或土壤中,毛細力很弱,邊緣變窄。
在毛細管邊緣和土壤表面之間是滲河流區或通風區。它含有空氣和微生物呼吸氣體,毛細水和水,通過重力向下移動到潛水區。 Pellicular 水是地下水的薄膜,附著在地下水位以上的單個顆粒上。這種水通過分子吸引力保持在毛細管邊緣上方。如果潛水區提供的水連續供應至孔,它被稱為一個含水層。良好的含水層通常具有相對於從井中抽出的水量和高孔隙度的大的橫向和垂直範圍,這允許水容易地排出。
含水層的另一面是透水層或封閉床。弱透水層或限制層是相對較薄的沉積物或岩石層,具有低滲透性。通過透水層的垂直水運動受到嚴格限制。如果一含水沒有限制層覆蓋它,它被稱為一個非承壓含水層。甲承壓含水層是一個由弱透水層限制。
含水層和隔水層的複雜性和多樣性導致了大量的含水層和隔水層下情景。例如,當有限大小的淺水層防止水向下移動到潛水區時,會發生棲息地下水。水收集在透水層上方並形成 � 小型潛水區 � 。在許多情況下,棲息的地下水僅在暴風雨期間或在潮濕季節出現。在旱季,井水可能會遇到缺水。但是,棲息含水層可能是當地重要的地下水源。
自流井是在承壓含水層中開發的。由於承壓含水層中的靜水壓力大於大氣壓力,因此自河流井中的水位上升到大氣壓等於靜水壓力的水平。如果該高度高於地面,則水可以自由地河流出井。
在地面與承壓含水層相交的地方,水也會自由河流動。測壓表面是其中水將上升在挖掘到承壓含水層的孔,如果井延伸無限期地表面之上的水平。潛水井從潛水區下方承壓含水層取水。潛水井中的水位與地下水位相同。
河道修復的我們應關注地下水和地表水交換的地點。該區域允許 fteely 水運動的潛水區被稱為補給區。地下水位與土壤表面相遇或河流水和地下水出現的區域稱為泉水或滲漏。
地下水量和地下水位的高低根據地下水的補給和排放而波動。由於水位升高的波動,一條小溪河道可以作為補給區域 ( 河流入或 � 河流失 � 河流 ) 或排放區域 ( 河流出物或 � 獲得 � 河流 ) 。

圖 2.8: 地下水相關用語
2A-1.3 逕流
當降雨或融雪的速度超過滲透能力時,多餘的水會聚集在土壤表面並作為逕流向下行進。影響逕流過程的因素包括氣候,地質,地形,土壤特徵和植被。美國毗鄰地區的年平均逕流範圍從不到 1 英寸到超過 20 英寸 ( 圖 2.9) 。
本小節介紹了三種基本類型的逕流 :
■ 地表逕流
■ 地下水流動
■ 飽和水中間河流
這些逕流類型中的每一種都可以單獨出現或在同一區域內以某種組合出現。
地表逕流
當降雨率超過滲透率時,水會在小凹陷中聚集在土壤表面 。 存儲在這些空間中的水被稱為抑鬱症儲存。它最終通過蒸發返回大氣或滲入土壤表面。凹陷的存儲空間被填充後,過量的水開始移動下坡路為坡面河流,無論是作為淺片的水或作為一系列小河流或細溝的。 Horton(1933) 是第一個在文獻中描述這一過程的人。通常使用術語 Horton 陸地河流或 Hortonian 河流。當水面向下移動時,水的深度和速度會增加。在旅行時,一些陸上河流動被困在山坡上,被稱為地表滯留。與向大氣蒸發或進入土壤的窪地儲存不同,地表滯留僅暫時停留在其下坡行程中。它最終河流入河流,仍然被認為是陸地河流量總量的一部分。
陸上河流動通常發生在具有鋪砌和不可滲透表面的城市和郊區環境中。已經被重型設備或車輛暴露和壓實的鋪砌區域和土壤也是陸上河流動的主要設置。在具有稀疏植被覆蓋的薄土壤區域中也常見,例如在乾旱或半乾旱地區的山區。
地下水流動
一旦進入土壤,水就會響應水頭的差異而移動 ( 由於不同高度的靜水壓力梯度導致河流動的可能性 ) 。 鑑於情況簡化,暴雨前的地下水位有一個拋向水面的拋物面。水沿著該斜坡向下移動並進入河流廊道。這部分河流程是基河流。地下水位下的土壤當然是飽和的。假 設山坡具有均勻的土壤特性,表面土壤的含水量隨著距河流的距離而減小。在暴雨期間,與土壤上坡相比,最靠近河流的土壤具有兩個重要屬性 - 較高的水分含量和較短的距離地下水位。這些屬性導致地下水位響應雨水滲透而更快地上升並導致地下水位變陡。因此,一種新的暴雨產生的地下水成分被添加到基河流中。這種稱為次表面河流動的新組件與基河流混合,增加地下水排放到河道。 在某些情況下,由於存在水層,滲透的雨水不會到達潛水區。在這種情況下,地下水流不會與基河流混合,但也會將水排放到河道中。無論是否混合,最終結果是增加了河道河流量。

圖 2.9: 台灣地區的平均年逕流量。但多為颱風季節迅速入海,水庫能截留量僅佔 6% 。

圖 2.10: 降雨後的水流路徑。河流到地下水位或滲入地下水位的降水量 , 取決於土壤的滲透率 ; 地表粗糙度 ; 以及降雨的數量、持續時間和強度

圖 2.11: 陸域逕流流動和凹陷儲存。地表逕流以不規則的流速向下移動。
飽和水與中間流
如果上述暴雨繼續,地下水位表面的坡度可能會繼續在河流附近變陡。最終,它可能會變得陡峭到水位高於河道高度的程度。此外,地下水可以從土壤中分離出來並作為陸地河流動進入河流。這種類型的逕流稱為 快速回滲流 。
地下水突破下面的土壤當然是飽和的。因此,達到最大滲透率,並且落在其上的所有降雨作為地表水流往下坡。這種直接降雨和快速流往河流的結合被稱為飽和的陸地逕流量。隨著暴雨的進展,飽和區域進一步延伸至山坡上。由於 快速回滲流 和地下水流與地表逕流流動密切相關,因此它們通常被認為是地表水總體逕流的一部分。
2A 河道中的水文水理過程
河流入河流的水是直接降雨和水從陸地橫向移動到河道中的集合。橫向運動的數量和時間直接影響
河流量的數量和時間,反過來影響河道的生態功能。
2A-1 水流分析
河流量範圍從無河流量到各種時間尺度的洪水流量。在廣泛的範圍內,歷史氣候記錄顯示偶爾持續的干濕年份。例如,美國的許多河流在 20 世紀 30 年代的 � 沙塵暴 � 十年中河流量下降。全國範圍內河流量的另一次類似下降發生在 20 世紀 50 年代。不幸的是,關於乾濕年的記錄時間很短 ( 在地質時間內 ) ,因此很難預測乾濕年的廣泛持續性。
河流量的季節性變化更具可預測性,但持久性因素有些複雜化。由於設計工作需要使用歷史信息 ( 記錄期 ) 作為未來設計的基礎,因此河流信息通常以概率格式呈現。兩種格式對規劃河廊復育特別有用:
■ 河流動持續時間,給定河流量在一段時間內等於或超過的概率。
■ 河流量頻率,給定河流量的概率將在一年內超過 ( 或不超過 ) 。 ( 有時這個概念被修改並表示為超過 [ 或不超過 ] 給定河流量之間的平均年數。 )

圖 2.12 降雨概率曲線與河流水流量關係的示例。該圖表顯示 x 軸上的月份和 y 軸上的每月平均水流範圍。曲線表示平均每月排放量小於曲線所示值的概率。例如,在 1 月 1 日左右,有 90 %的可能性排放量將低於 9,000cms , 50 %的可能性將低於 2,000cms 。
2A-2 水流的生態影響
河流量的變化是決定河流生態系統結構和動態的生物和非生物過程的主要影響 (Covich1993) 。高河流量不僅在沉積物運輸方面很重要,而且在將洪氾平原濕地重新連接到河道方面也很重要。
這種關係非常重要,因為洪氾平原濕地為魚類提供了產卵和育苗棲息地,並在今年晚些時候為水禽覓食棲息地。低河流量,特別是在大型河流中,創造條件,允許支流動物群分散,從而維持
在幾個地點的單一物種的種群。
一般而言,許多河流物種的生命週期的完成需要一系列不同的棲息地類型,其時間可用性由河流動狀態決定。適應這種環境活力使得河流物種能夠在乾旱和洪水期間持續存在,從而破壞和重建棲息地元素。
2B 地貌過程
地貌學是研究地球表面形態和形成這些形態的過程。上一節中討論的水文過程驅動了本節中描述的地貌過程。反過來,地貌過程是形成第 1 章討論的排水模式,河道,洪氾區,梯田和其他河流流域以及河道特徵的主要機制。
河流水涉及三個主要的地貌過程,如下:
■ 侵蝕,土壤顆粒的分離。
■ 泥砂運輸,河流動的水中侵蝕的土壤顆粒的運動。
■ 沉積物沉積,侵蝕的土壤顆粒沉澱到水體底部或隨著水分離開而留下。
沉積物沉積可以是短暫的,如在從一個暴雨到另一個暴雨的河流廊道中,或者或多或少永久性,如在較大的儲層中。
由於地貌過程與水的運動密切相關,因此本節被組織成子區域,反映了地表雨水逕流和河流的水文過程:
■ 河道橫向的地貌過程
■ 河道縱向的地貌過程
2B-1 整個河流的地貌過程
河流流域侵蝕過程的發生,大小和分佈會影響河道的沉積物和相關水質污染物的產量。
土壤侵蝕可以在很長一段時間內逐漸發生,或者可以是循環的或偶發的,在某些季節或某些暴雨事件期間加速 ( 圖 2.13) 。土壤侵蝕可能是由人類活動或自然過程引起的。侵蝕不是一個簡單的過程,因為土壤條件隨著溫度,水分含量,生長階段和植被數量以及人類對土壤的操縱而不斷變化,以促進開發或作物生產。表 2.2 和表 2.3 顯示了影響土壤侵蝕的基本過程以及河流流域內發現的不同類型的侵蝕。
2B-2 河流中的地貌過程
河道,洪氾區,平台,和其他在該河道特徵形成主要通過侵蝕,運輸,和沉積物的通過沉積逕流。本小節描述了向下游輸送沉積物承載以及河道和洪氾區如何隨時間調整和演變所涉及的過程。
2B-2.1 泥砂搬運
在河道和洪氾區中發現的沉積物顆粒可根據大小進行分類。巨石是最大的顆粒,粘土是最小的顆粒。顆粒密度取決於顆粒的尺寸和組成 ( 即顆粒的礦物質含量的比重 ) 。
無論大小如何,河道中的所有顆粒都可以在下坡或下游運輸。最大顆粒上的河流可以在給定的水理條件下移動的大小被稱為為河流能力。通常,只有非常高的河流量能夠移動最大的顆粒。
與水流能力密切相關的是牽引應力的概念,它在沿河床和河岸的河流邊界處產生升力和阻力。牽引應力,也稱為剪切應力,隨河流動深度和坡度而變化。假設密度,形狀和表面粗糙度恆定,顆粒越大,移動它並將其向下游移動所需的牽引應力越大。
將沉積物顆粒置於運動中的能量來源於更快的水流過較慢的水的影響。發生這種速度梯度是因為河流體主體中的水比邊界處河流動的水移動得更快。
如果河床是粗糙的,當河流動在它們上方時產生摩擦,這反過來會減緩河流動。
較快水的動量傳遞到較慢的邊界水。在這樣做時,較快的水傾向於以螺旋運動將較慢的水捲起。正是這種剪切運動或剪切應力也使床顆粒在下游的滾動運動中移動。
河道底部的粒子運動以滑動或滾動運動開始,沿著河流動方向沿著河床傳輸粒子 ( 圖 2.14) 。一些顆粒還可以通過移動床表面上方跳躍,當一個顆粒與另一種粒子碰撞,導致其反彈向上,然後回落朝向床滿足跳躍運動。
這些滾動,滑動和跳躍運動導致移動的顆粒與河床的頻繁接觸,並且表徵被稱為床載荷的一組移動顆粒。這些顆粒相對於河流速的重量使它們在向下游移動時基本上保持與河床接觸並由河床支撐。
懸掛載荷的部分可以是膠態粘土,其可以留在懸浮液很長的時間週期,這取決於粘土和水化學的類型。
表 2.2: 土壤侵蝕過程。

表 2.3: 侵蝕類型與物理過程。


圖 2.13: 雨滴的影響。土壤侵蝕的多種類型之一。

圖 2.14: 水對河床附近顆粒的相互作用。運輸床載沉積物的過程是河流速、顆粒大小和水動力原理的功能。
沖刷承載與沉積承載
區分河流沉積物承載的一種方法是根據運輸中沉積物的直接來源對其進行表徵。在任何給定的時間和地點,河流中的總沉積物承載分為兩部分 - 洗滌承載和床料承載。洗滌承載的主要來源是河流流域,包括板塊和細溝侵蝕,溝壑侵蝕和上游河床侵蝕。床料承載的來源主要是河床本身,但包括河流流域中的其他來源。
洗滌承載由運輸中最細的沉積物顆粒組成。湍河流使洗滌承載保持懸浮狀態。懸浮液中洗滌承載的濃度基本上與河流中的水力條件無關,因此不能使用測量或估計的水力參數 ( 例如速度或排放 ) 來計算。洗滌承載濃度通常是供應的功能 ; 也就是說,該河流可以攜帶與河流流域和河岸可以輸送的一樣多的洗滌承載 ( 對於低於約百萬分之 3000 的沉積物濃度 ) 。
床料承載由河床中發現的尺寸等級的沉積物組成。床料承載通過滾動,滑動或跳躍沿著河床移動,並且可以通過湍河流週期性地夾帶到河流中,其中湍河流成為懸掛承載的一部分。床料承載是水理控制的,可以使用第 8 章討論的沉積物輸送公式計算。
泥砂搬運用語
泥砂搬運術語有時會令人困惑。由於這種混淆,定義一些更常用的用語很重要。
■ 沉積物承載量,指在一段特定時間內 ( 通常為一天或一年 ) 經過河流任何一個橫截面的沉積物數量。沉積物排放,沉積物的質量或體積在單位時間內通過河流橫截面。沉積物承載量的典型單位是噸,而沉積物排放單位是噸 / 每天。
■ 床料承載量,是由沉積物顆粒組成的總沉積物排放的一部分,沉積物顆粒與河床沉積物的大小相同。
■ 沖洗承量載,總沉積物承載的一部分,比河床中發現的顆粒尺寸更細的顆粒尺寸組成。
■ 河床承載量,在河床層中通過鹽化,滾動或滑動在河床上或附近移動的總沉積物承載的一部分。
■ 懸浮床物質承載量,在水體中懸浮運輸的床料承載的一部分。懸浮床物質承載和床承載總加,為總河床物質承載量。
■ 懸浮物質承載量,被輸送的總沉積物承載的一部分,通過水流驅動的主要懸浮物質。
■ 測量承載量,採樣區採樣器獲得的總沉積物承載的一部分。
■ 未測量的承載量,即懸浮水體中和河床上通過取樣器下方的總沉積物承載的一部分。對於典型的懸浮沉積物採樣器,這是垂直的 0.1 至 0.12 公尺。
上述術語可以通過多種方式組合,以確定河流中的總沉積物承載 ( 表 2.4) 。但是,重要的是不要組合不兼容的術語。例如,懸掛載荷和床料載荷不是互補條款,因為懸浮載荷可包括床料載荷的一部分,這取決於可用於運輸的能量。總沉積物承載通過以下術語的組合正確定義:
總沉積物承載 =
床料承載 + 洗滌承載
或
床承載 + 懸掛承載
或 測量承載 + 未測量承載沉積物輸送速率可使用各種公式或模型計算。
這些將在第 8 章的河流河道復育部分中討論。
表 2.4: 沉積物承載量分項

水流動力
水流的主要地貌任務之一是將粒子輸送出河流流域。以這種方式,河流用作運輸 � 機器 � ,並且作為機器,其工作速率可以被計算為可用功率乘以效率的乘積。河流功率可以計算為:
ψ = γ Q S
ψ= 河流動力 (foot-lbs/second-foot 作用點通過壓力時通過一磅力進行的功 / 每秒經過指定點的一立方英尺的水流量 ) ; ( 公尺 - 千克 / 秒 -M 3 )
γ = 水的比重 (lbs/ft 3 )(kg/M 3 )
Q= 水流 (ft 3 /second)(M 3 / 秒 )
S = 斜率 (feet/feet)(M/M)
泥砂輸送速率與河流動力直接相關 ; 即坡度和河流量。基河流下面的高度彎曲的河流谷底線以曲折河流 ( 即沿河流河道標記的最深點的線 ) 幾乎不產生河流功率 ; 因 此,河流移動沉積物,沉積物運輸能力的能力是有限的。在更深的地方,河流動遵循更直的路線,這增加了斜坡,導致沉積物運輸速率增加。該河流建立其橫截面以 獲得河流動深度和河道斜率,其產生維持河流河道所需的沉積物輸送能力。由於自然原因或土地,逕流可能因河流流域而異使用實踐。這些變化可以通過優先將特定 的顆粒尺寸移動到河流中來改變從河流流域輸送到河流中的沉積物的尺寸分佈。這是並不少見到鵝卵石層的頂部找到沙層。這通常發生在沙質土壤在河流流域中發生 加速侵蝕時,並且在將沙子移入河道的事件期間,增加的沙子承載超過了河流的運輸能力。

圖 2.15: 土砂粒子傳輸。河流沉積物負荷是指定時間段內通過特定橫截面的沉積物顆粒總和。輸砂能力因運輸機制而異。
2B-2.2 河流與洪氾區穩定性
在考慮任何河流復育動作時通常會出現的一個問題是 � 現在它是否穩定並且在進行更改後它是否會穩定? � 答案可能比較基於捲軸中的幾幀來詢問對電影的意見。雖然我們經常根據時間的有限參考來查看河流,但重要的是我們要考慮河道橫截面,縱向輪廓和平面形態的長期變化和趨勢來表徵河道穩定性。
實現河道穩定需要的平均牽引應力保持穩定的河床和河岸坡。那需要河流的每個部分中的顆粒尺寸分佈保持平衡 ( 即,沉積的新顆粒與通過牽引移位的顆粒具有相同的尺寸和形狀強調 ) 。楊 (1971) 改編了利奧波德所描述的基本理論來解釋河流的縱向剖面,河流網絡的形成,湍瀨,深潭和河流蜿蜒。所有這些河流特徵和沉積物運輸密切相關。 Yang(1971) 基於熵概念,發展了平均河流量下降理論和最小能量消耗率理論。這些理論表明,在向平衡條件演化過程中,自然河流選擇其河流動過程,使得沿其路線的每單位質量河流量的潛在能量消耗率最小。
河道調整
河流河道及其洪氾平原不斷適應河流流域提供的水和沉積物。成功復育退化河流需要了解河流流域歷史,包括自然事件和土地利用實踐,以及在河道演變中積極的調整過程。
河道對水和沉積物產量變化的響應可能發生在不同的時間和地點,需要不同程度的能源消耗。河流量和沉積物承載的每日變化導致在具有可移動床的許多河流中頻繁 調節床形和粗糙度。河流還定期調整至極端高地低河流量事件,因為洪水不僅消除了植被,而且沿著河道產生並增加了植被潛力 ( 例如,低河流量時期允許植被侵入河道 ) 。
河流入廊道和高地河流流域的土地利用變化也可能帶來類似的調整。同樣,氣候變化,野火等自然原因導致的逕流或沉積物產量的長期變化,或人為原因,如種植, 過度草食者或從農村到城市的轉變,都可能導致長期調整在河道橫截面和平面圖中,經常被描述為河道演變。
在許多研究中已經定性地描述了河流廊道對河流量和沉積物承載變化的響應 ( 例如, Lane1955 , Schumm1977) 。如第 1 章所述, Lane(1955) 提出了解釋河流行為的最早關係之一,他將平均年河流量 (Q W ) 和河道坡度 (S) 與河床沉積物相關聯。
河床上的載荷 (Qs) 和中值粒徑 (D 50 ) :
Q 5 � D 50 〜 Q W � S
Lane 的關係表明,當沉積物承載和床料大小的變化通過河流量或河道梯度的變化來平衡時,河道將保持動態平衡。這些變量之一的變化引起一個或多個其他變量的變化,從而重新建立動態平衡。
已經提出了額外的定性關係來解釋沖積河道的行為。 Schumm(1977) 認為寬度 (b) ,深度 (d) 和曲折波長 (L) 是成正比的,並且河道梯度 (S) 與河流量成反比 ( 沖積河道中的
Q W 〜 b,d,L /S
Schumm(1977) 也提出寬度 (b) ,曲折波長 (L) 和河道梯度 (S) 成正比,深度 (d) 和彎曲度 (P) 與沉積物水流 (Qs) 成反比。沖積河流:
Q S 〜 b,d,L / d,P
考慮到河流量或沉積物排放的增加或減少,可以重寫上述兩個公式以預測河道特性的變化方向:

將上述四個公式組合起來,可以為河流量和 / 或沉積物排放的同時增加或減少提供額外的預測關係:

河道坡降
河道斜率 ( 河流的縱向輪廓 ) 被測量為河流中兩點之間的高度差除以兩點之間的河流長度。斜率是考慮河道修改時所需的最重要的設計信息之一。河道斜率直接影響河流速,河流能力和河流動功率。由於這些屬性驅動了侵蝕,沉積物輸送和沉積物沉積的地貌過程,因此河道坡度成為河道形狀和模式的控制因素。
河流的大多數縱向剖面在上游是下凹的。如先前在動態平衡的討論中所述,河流調整它們的輪廓和模式以試圖最小化河流動水中存在的勢能或河流動力的時間消耗 率。河流的輪廓的凹形向上形狀似乎是由於河流的調整,以幫助最小化下游方向的河流動力。 Yang(1983) 應用最小河流動力理論來解釋為什麼大多數縱向河床剖面是向上凹的。為了滿足最小河流動力理論,這是一般最小理論的一個特例。
能量耗散率 (YangandSong1979) ,必須滿足以下等式:

其中:
P=QS= 河流功率
x= 縱向距離
Q= 排水量
S= 水面或能量斜率
γ = 水的比重
河流動功率已被定義為水流和斜率的乘積。由於河流水流通常在下游方向上增加,因此必須減小斜率以使河流功率最小化。下游方向上的斜率的減小導致凹陷的縱向輪廓。 Sinuosity 不是輪廓特徵,但它確實影響河流坡降。彎曲度是河流上兩點之間的河流長度除以兩點之間的谷長度。例如,如果從 A 點到 B 點的河流長度為 2,200 英尺,並且如果這兩個點之間的谷長距離為 1,000 英尺,則該河流的彎曲度為 2.2 。河流可以通過增加其彎曲度來增加其長度,從而導致斜率減小。這種彎曲對河道坡度的影響如果河道重建是擬議修復的一部分,則必須始終考慮。
河流動功率已被定義為水流和斜率的乘積。由於河流水流通常在下游方向上增加,因此必須減小斜率以使河流功率最小化。下游方向上的斜率的減小導致凹陷的縱向輪廓。 Sinuosity 不是輪廓特徵,但它確實影響河流坡降。彎曲度是河流上兩點之間的河流長度除以兩點之間的谷長度。例如,如果從 A 點到 B 點的河流長度為 2,200 英尺,並且如果這兩個點之間的谷長距離為 1,000 英尺,則該河流的彎曲度為 2.2 。河流可以通過增加其彎曲度來增加其長度,從而導致斜率減小。這種彎曲對河道坡度的影響如果河道重建是擬議修復的一部分,則必須始終考慮。
深潭與湍瀨
即 使在短距離內,縱向輪廓也很少是恆定的。地質,植被模式或人為干擾的差異可導致整體輪廓內的更平坦和更陡峭的河段。湍瀨發生瓦特這裡的河流底部是相對更高 海拔河床立即上游或下游。這些相對較深的區域被認為是水潭。在正常河流動時,水潭面積中的河流速降低,允許發生細粒度沉積,並且由於湍瀨頂部和隨後的水潭 之間的床坡度增加而增加頂部湍瀨。
縱向剖面調整
當在河流上構建壩時,發生輪廓調整的常見示例。對大壩建設的典型反應是下游的河道退化和上游的加速。但是,具體的響應非常複雜,可以通過考慮 Lane 的關係來說明。大壩通常會減少下游河段的峰值排放和沉積物供應。根據 Lane 的關係,排放量的減少 (Q) 應該通過斜率的增加來抵消,但是沉積物承載的減少 (Qs) 應該導致斜率的減少。如果發生裝甲 (D^t) ,這種反應可能會進一步複雜化,這也會導致斜率增加。影響不僅限於主要河道,還可以包括支流的加重或退化也是如此。由於主幹上的峰值河流量減少,通常在大壩下游 ( 有時在整個河道 ) 的支流口處發生沉積。顯然,最終的反應將是所有這些變量整合的結果。
河道橫斷面
圖 2.16 顯示了收集河流截面數據時應記錄的信息類型。在穩定的沖積河流中,每個河岸的高點代表了滿槽河道的頂部。 已經建立了水岸河道的重要性。河道橫截面需要包括足夠的點以定義與每一側的氾濫平原的一部分相關的河道。建議的指南是在較小的河道上至少包括一個超出每個河岸最高點的河流寬度,並且在較大的河流上至少包括足夠的洪氾區以清楚地定義其相對於河道的特徵。
在 曲河流中,應在淺溝或交叉區域測量河道橫截面。在兩個連續曲折的頂點之間發生淺溝或交叉。在曲折的外側彎曲處,土壤層的抗侵蝕性的差異的影響是顯著的,並 且曲折的內側上的點條不斷地調整到由河流移動的水和沉積物承載。河流的橫截面在曲折彎曲中變化得更快更頻繁。水潭橫截面的變化比淺槽橫截面的變化更大。交 叉或淺溝區域的橫截面更均勻。

圖 2.16: 河道斷面。收集河流橫斷面資料時要記錄的資訊。
抗河流動性與河流速
河道斜率是確定河流量河流速的重要因素。河流速用於幫助預測橫截面可以傳達的排放。隨著排放增加,河流速,河流動面積或兩者都必須增加。
粗糙度在河流中起著重要的作用。它有助於確定河流範圍內的河流動深度或階段。由於粗糙度導致河流速減慢,河流動速度必須增加,以保持進入河段範圍上游端的河流量 ( 稱為河流動連續性的概念 ) 。沿著河流的邊界的典型粗糙度包括以下:
■ 不同大小的沉積物顆粒。
■ 床型。
■ 水岸違規行為。
■ 生命和死亡植被的類型,數量和分佈。
■ 其他障礙物。
粗糙度通常隨著粒度的增加而增加。的形狀和尺寸插播沉澱沉積物或底形,也有助於粗糙度。砂底河流是床形粗糙度隨排放變化的良好例子。在非常低的排放下,砂 河流床可以由波紋床形狀支配。隨著河流量的增加,沙丘可能開始出現在床上。這些床形中的每一個都增加了河流底部的粗糙度,這傾向於減慢速度。
由於粗糙度增加,河流動深度也增加。如果水流繼續增加,則達到一個點當河流速在河床上移動沙子並且整個床再次轉換成平面形式時。由於床的粗糙度降低,此時 河流動的深度實際上可能減小。如果水流仍然進一步增加,則可能形成抗結核。這些床型產生足夠的摩擦力,再次導致河流動深度增加。河流在砂床河流的給定的水 流深度,因此,依賴於底形當水流發生時出現。植被也可能導致粗糙。在具有由粘性土壤組成的邊界的河流中,植被通常是粗糙度的主要成分。
河道中植被的類型和分佈取決於水文和地貌過程,但通過創造粗糙度,植被可以改變這些過程並引起河流形態和模式的變化。蜿蜒的河流相對於直河流提供一些河流動阻力。直的和曲折的河流也具有受影響的不同河流速分佈通過河流的對齊,如圖 2.17 所示。在河流的直線範圍內,最快的河流動發生在河道中心附近的表面下方,其中河流動阻力最低 ( 見圖 2.17(a)G 部分 ) 。在曲折中,由於角動量,外邊緣的速度最高 ( 見圖 2.17(b) 第 3 節 ) 。
曲折河流中河流速分佈的差異導致曲折彎曲處的侵蝕和沉積。侵蝕發生在彎曲部 ( 的外部 cutbanks 從高速河流 ) ,而在彎曲部的內側較慢速度導致點欄上沉積 ( 其也被稱為在脫斜率 ) 。通過曲折彎曲的角動量增加了高度或在彎曲外側的超高度,並設置一個次要的河流動,沿著切割堤的面向下河流動,越過深潭底部朝向彎曲內側。這個旋轉河流被稱為螺旋 flow.and 是通過在橫截面 3 的在圖中的頂部和底部和 4 中的箭頭所示的圖上的下頁的旋轉方向。
直河流和曲河流中的河流速分佈對於在河流路廊修復中規劃河流對齊的修改時非常重要。最高速度的區域產生最大的河流動力,因此在這樣的速度與河流動邊界相交 的地方表明可能需要更持久的保護。當河流動通過曲折時,水潭中的底部水和碎屑旋轉到表面。這種旋轉是移動漂浮和底棲生物過去水潭中捕食者的重要機制。
湍瀨區不像水潭區那麼深,因此在這些淺層區域中會出現更多的湍河流。湍瀨流可以增加水的溶氧含量,也可以增加水中某些化學成分的氧化和揮發。粗糙元素的另一個非常重要的功能是它們創造了水生棲息地。作為一個例子,最深的河流動深度通常發生在切割庫的底部。與滑脫斜坡的沉積環境中發生的相比,這些沖刷洞或水潭形成非常不同的棲息地。

圖 2.17: (a) 直河流分支和 (b) 河流蜿蜒。河流河流速度是不同的 , 通過水潭和湍瀨 , 在直和彎曲的河段 , 跨越河流在任何點 , 在不同的深度。速度分佈也與通過河岸通河流和洪水流的基本河流量分佈有顯著差異。 Source: Leopold et al. 1964. Published by permission of Dover Publications.

圖 2.17a: 廣義速度分佈
2B-3 主深槽與洪氾區
洪氾區由兩個河流程構建,即橫向和縱向增生。側向增生是沉積物沉積在河流彎道內側的點棒上。當河曲彎曲的外部侵蝕並且點條形成有粗糙紋理的沉積物時,河流橫向遷移穿過洪氾平原。這種自然發生的過程保持了輸送水和水的橫截面
來自河流流域的沉積物。垂直吸積是沉積物在淹水表面上的沉積。這種沉積物通常比點狀沉積物具有更細的紋理,並且被認為是過岸沉積物。垂直吸積發生在點棒的側向吸積沉積物之上 ; 然而,橫向吸積是主要的過程。它通常佔洪氾區沉積物總沉積量的 60 %至 80 % (Leopold 等, 1964) 。很明顯,蜿蜒的橫向遷移是一個重要的自然過程,因為它在重塑氾濫平原方面起著關鍵作用。

圖 2.18: 河流沉降。雖然沉積物及其運輸是自然發生的 , 但沉積物載荷和顆粒大小的變化會產生負面影響。
2C 物理和化學特性
河道中的水質可能是復育的主要目標,要么將其改善到理想狀態,要么維持它。如果水的物理和化學特性不合適,在河道中建立適當的河流態和地貌可能對確保健康 的生態系統沒有多大幫助。例如,含有高濃度有毒物質或其中高溫,低溶氧或其他物理 / 化學特性不合適的河流不能支持健康的河道。相反,河道的狀況不佳 - 例如缺乏河岸遮蔭,對侵蝕的控制不良,或過多的養分來源和氧氣需求廢物會導致物河流內物理和化學條件的降低。
本節簡要介紹了河流動水的一些關鍵物理和化學特徵。河流水質是一個廣泛的主題,許多書籍都寫在這個主題上。這裡的重點是
一個是相關的幾個關鍵概念,河道復育。讀者可以參考其他來源 ( 例如, Thomann 和 Mueller1987 , Mills 等人 1985) 進行更詳細的治理。
如前面部分所述,在橫向和縱向視角中檢查了物河流的物理和化學特性。橫向視角指的是河流流域對水質的影響,特別是對河岸地區的關注。縱向透視是指在運輸河道中影響水質的過程。
2C-1 物理特性
2C-1.1 沉積物
第 2.B 節 討論了在河流形態和地貌演化的背景下的總沉積物承載。除了它在塑造河流形態中的作用外,懸浮沉積物在水體和沉積物水界面的水質中起著重要作用。在水質環境 中,沉積物通常是指從侵蝕的土地進入水體的土壤顆粒。沉積物由各種尺寸的顆粒組成,包括細粘土顆粒,淤泥和礫石。術語沉降用於描述沉積物顆粒在水體中的沉 積。
雖然沉積物及其運輸在任何河流中自然發生,但沉積物承載和粒徑的變化可能產生負面影響 ( 圖 2.18) 。 細小的沉積物會嚴重改變水生生物群落。沉積物可能會堵塞和磨損魚鰓,在底部窒息雞蛋和水生昆蟲幼蟲,並填充魚卵產卵的底部鵝卵石之間的孔隙空間。沉積物通 過降低水的透明度和填充水體來干擾水體的遊憩活動和美學享受。沉積物也可能將其他污染物帶入水體。營養物質和有毒化學物質可能附著在陸地上的沉積物顆粒 中,並將顆粒帶入地表水中,污染物可能會沉積在沉積物中或變得可溶於水體中。
研究表明,細小的沉積物侵入可以顯著影響產卵棲息地的質量 (Cooper1965 , Chapman1988) 。細顆粒泥砂侵入河床礫石可以減少透氣性和 intragravel 水流速度,從而限制的含氧水開發供應鮭魚胚胎和去除它們的代謝廢物。過量的細小沉積物沉積可以有效地扼殺孵化蛋和孵化 alevins 和魚苗。沉積物侵入模型 (Alonso 等人, 1996) 已被開發,驗證和驗證,以預測批內里德 ( 產卵區 ) 沉積物堆積和溶氧狀態。
整體河流的沉積物
雨水侵蝕並將犁過的田地,建築工地,伐木場所,城市地區和帶狀土地上的土壤顆粒沖洗到水體中。侵蝕河岸坡還存泥砂進入水體。總之,河流中的沉積物質量代表了河流流域侵蝕過程的最終結果。
沉積物的側視圖在第 2.B 節中有更詳細的討論。然而,值得注意的是,從水質角度來看,興趣可能集中在沉積物承載的特定部分。例如,控制細小的沉積物承載通常是復育鮭科魚類棲息地的特別關注點。
復育工作可用於控制從河流流域到河流的沉積物和沉積物相關污染物的承載。這些可能包括減少高地侵蝕的努力,以及減少通過河岸帶的沉積物輸送的處理。第 8 章介紹了修復處理的設計。
縱向各河段的沉積物
從水質角度影響沉積物輸送的縱向過程與第 2.B 節中從地貌角度討論的過程相同。如在側向視角中,從水質觀點來看,由於其對水質,水溫,棲息地和生物群的影響,從水質觀點出發的興趣可能集中在特定的沉積物大小部分,特別是細沉積物部分。
2C-1.2 水溫
由 於多種原因,水溫是河道復育的關鍵因素。首先,溶氧的溶解度隨著水溫的升高而降低,因此需氧廢物所施加的應力隨著溫度的升高而增加。其次,溫度控制著冷血 水生生物中的許多生化和生理過程,溫度升高可以提高整個食物鏈的代謝和繁殖率。第三,許多水生物種只能承受有限的溫度範圍,並且改變河流內的最高和最低溫 度會對物種組成產生深遠的影響。最後,溫度也影響許多非生物化學過程,例如再氧化速率,有機化學物質吸附到顆粒物質和揮發率。溫度升高可導致有毒化合物的 應力增加,其中溶解的部分通常是最俱生物活性的部分。
整個河流的水溫
河 段的河流入水溫度的最重要因素是通過地面和地下水路徑到達的水之間的平衡。河流過陸地表面的水流有機會通過與太陽加熱的表面接觸而獲得熱量。相比之下,夏 季地下水通常較涼,並且往往反映了河流流域的年平均溫度。通過淺地下水河道的水流可能位於逕流事件期間的年平均溫度和環境溫度之間。兩個逕流經由表面途徑 和地表逕流的溫度到達分數被強烈影響通過河流流域內不透水表面的數量。例如,河流流域中的熱鋪面可以加熱地表逕流,並顯著提高接收逕流的河流溫度。
河道水體的水溫
水也會受到陽光直接影響河流的熱承載。對於復育的目的, � 土地使用做法除去開銷蓋或減少 baseflows 可以增加河道內溫度超過了魚類臨界火最大值的水平 (Feminella 和馬修斯 1984) 。因此,維持或復育正常溫度範圍可以是復育的重要目標。前面的章節討論了水在水文循環中的物理旅程。雨水滲透到地下水位或變成陸上河流動,溪水收集這些水並將其引向地下水。

圖 2.19: 溪流懸浮沉積物上的有機塗層。水化學確定沉積物是攜帶吸附材料 , 還是對溪流沉積物進行塗層。
2C-2 化學特性
海洋和蒸發蒸騰在整個循環中發生。隨著水的到來,它的化學變化。在空氣中,水與大氣氣體平衡。在 .shallow 土壤中,它與無機和有機物質以及土壤氣體進行化學交換。在地下水中,運輸時間較長,礦物質溶解的機會更多。類似的化學反應沿著河道繼續進行。每到一處,水相互作用 eveiything 倒是空氣,岩石,細菌,植物和魚類,並通過人為干擾的影響。
科學家已經能夠為水中許多常見的溶解成分定義幾個相互依賴的循環。這些循環中的中心是氧,碳和營養素的行為,例如氮 (N) ,磷 (P) ,硫 (S) 和較少量的常見微量元素。
例如,鐵是動植物新陳代謝的必需元素。水生系統中的鐵可以以兩種氧化態之一存在。三價鐵 (Fe3+) 是更加氧化的形式,並且非常微溶於水。還原形式的亞鐵 (Fe2+) 更易溶解許多個數量級。在許多水生系統中,例如湖泊,鐵可以從鐵狀態循環到亞鐵狀態並再次返回 ( 圖 2.19) 。 亞鐵的氧化隨後三價鐵的沉澱導致一些河流沉積物表面上的鐵塗層。這些塗料以及有機塗料在有毒微量元素和有毒有機化學品的水生化學中發揮著重要作用。有毒有機化學品和金屬的化學以及氧,氮和磷的循環和化學將在本節後面介紹。
水中所有溶解離子的總濃度 ( 也稱為鹽度 ) 變化很大。沉澱通常僅含有千分之幾 (ppt) 的溶解固體,而海水的鹽度平均約為 35ppt( 表 2.5) 。淡水中溶解固體的濃度可能在原始山區河流中僅為 10 至 20mg/L ,在許多河流中為幾百 mg/L. 乾旱河流流域的濃度可能超過 1,000 毫克 / 升。小於 500 毫克 / 升的溶解固體濃度建議對於公共飲用水,但在該國許多地區超過了這一門檻。一些作物 ( 特別是果樹和豆類 ) 對即使是適度的鹽度也很敏感,而其他作物,如棉花,大麥和甜菜,可以耐受高濃度的溶解固體。灌溉產生的農業回水可能會增加河流的鹽度,特別是在西部。建議的家畜鹽度限制從家禽的 2,860 毫克 / 升到成年綿羊的 12,900 毫克 / 升不等。植物,魚類和其他水生生物也 vaiy 的適應廣泛,以不同濃度溶解固體。大多數物種有一個最大的耐鹽性及少數可生活在非常純淨水 veiy 低離子濃度。
表 2.5: 降雨的化學成分 (mg/ 公升 ) 。

1. 內華達州山頂的降雪。 2. 北卡羅來納州的降雨。 3. 加州 1 月的降雨。 4. 加州 1 月的降雨。 5. 美國內陸的降雨採樣。 6. 華盛頓州平均降水組成。
2C-2.1 pH 值 - 酸鹼度
鹼度,酸度和緩衝能力是水的重要特徵,影響其對生物群的適應性並影響化學反應。水的酸性或鹼性 ( 鹼性 ) 通常通過氫離子濃度或 pH 的負對數來量化。 pH 值為 7 表示中性條件 ;pH 值小於 5 表示中等酸性條件 ;pH 值大於 9 表示中等鹼性條件。許多生物過程,例如繁殖,不能在酸性或鹼性水中起作用。特別是,在持續暴露於低 pH 值水域的情況下,水生生物可能遭受滲透不平衡。
快速 pH 波動也會對水生生物造成壓力。最後,酸性條件還會通過增加溶解度來加劇有毒污染問題,導致儲存在河流沉積物中的有毒化學物質的釋放。
整個河流的 pH 值
逕流的 pH 值反映了降雨和陸地表面的化學特徵。除了具有顯著海洋噴霧的區域外,大多數降雨中的主要離子是碳酸氫鹽 (HCO) 。碳酸氫根離子通過二氧化碳與水反應產生:

該反應還產生氫離子 (H+) ,從而增加氫離子濃度和酸度並降低 pH 。因為 CO 的存在的 2 在大氣中,雨最自然是微酸性的, pH 約 5.6 。降雨的酸度增加可能是由於投入,特別是燃燒化石燃料造成的。當水通過土壤和岩石移動時,其 pH 值可能隨著額外的化學反應而增加或減少。碳酸鹽緩衝系統控制大多數水的酸度。碳酸鹽緩衝由水中的鈣,碳酸鹽,碳酸氫鹽,二氧化碳和氫離子與大氣中的二氧化碳之間的化學平衡產生。緩衝導致水抵抗變化 pH 值。鹼度是指水的酸中和能力,通常是指那些使 pH 向鹼性方向移動的化合物。緩衝量與鹼度有關,主要由碳酸鹽和碳酸氫鹽濃度決定,碳酸鹽和碳酸氫鹽濃度由溶解的碳酸鈣 ( 即石灰石 ) 和類似物質引入水中。
河流流域中存在的礦物質。例如,當酸與石灰石相互作用時,會發生以下溶解反應:

該反應消耗氫離子,從而提高水的 pH 。相反,當水中的所有鹼度被酸消耗時,逕流可能會酸化,這一過程通常歸因於來自酸性礦山排水的強無機酸 ( 如硫酸 ) 和弱有機酸 ( 如腐殖酸和富裡酸 ) 的輸入,在某些類型的土壤中大量天然產生的土壤,例如與針葉林,沼澤和濕地有關的土壤。在某些河流中,通過復育干擾酸性輸入的退化濕地 ( 如酸性礦山排水 ) 可以提高 pH 值,並通過將硫酸鹽從硫酸轉化為不溶性非酸性物質來幫助中和酸度殘留在濕地沉積物中的金屬硫化物。
河流水體中的 pH 值
在河流中,在水中的酸,大氣 CO ,水體中的鹼度和河流線型物質之間發生類似的反應。在一些不良緩沖水中 pH 的另一個特徵是 pH 水平的高日變化可歸因於影響碳酸鹽緩衝系統的生物過程。在具有大型水生植物作物的水域中,植物在光合作用期間吸收二氧化碳會從水中去除碳酸,這會使 pH 增加幾個單位。相反,在夜間光合作用不發生且植物釋放出二氧化碳時, pH 值可能會下降幾個單位。增加遮蔭或減少養分含量來減少植物下游生長或增加耕種的恢復技術,也可以穩定由於高光合作用而引起的 pH 值高度可變的問題。
河流內的 pH 值可能對有毒物質產生重要影響。高酸度或高鹼度傾向於將不溶性金屬硫化物轉化為可溶形式並且可以增加有毒金屬的濃度。相反,高 pH 值可以促進氨毒性。氨以兩種形式存在於水中,即非離子化 (NH3) 和離子化 (NH/) 。在這兩種形式的氨中,非離子化氨對水生生物的毒性相對較高,而離子化氨的毒性相對可忽略不計。確定未離子化氨的比例通過水的 pH 和溫度 (Bowie 等人, 1985)- 當 pH 或溫度升高時,未離子化的氨的比例和毒性也增加。例如,當 pH 值為 7 且溫度為 20 蚓 時,只有約 0.4 %的總氨處於非離子化形式,而 pH 值為 8.5 且溫度為 25.56 蚓 時, 15 %的氨處於非離子化狀態。總氨,處於非離子化形式,對水生生物的潛在毒性增加 35 倍。
2C-2.2 溶氧
溶氧 (DO) 是健康水生生態系統的基本要求。大多數魚類和水生昆蟲 � 呼吸 � 溶解在水體中的氧氣。一些魚類和水生生物,如鯉魚和糞便蠕蟲,適應低氧條件,但如果 DO 濃度低於 3 至 4mg/L 的濃度,大多數活躍運動的魚類如馬口魚和粗首 鱲 都會受到影響。幼蟲和幼魚對氧的需求更敏感,需要更高濃度的 DO 。
許多魚類和其他水生生物可以從水中短時間的低 DO 中恢復過來,但是延長的溶氧濃度降低 2 毫克 / 升或更低的事件可導致魚類大量死亡。長時間接觸低 DO 條件將使魚類窒息,或因卵和幼魚的窒息降低其繁殖生存,或因水生昆蟲幼蟲大量死亡而失去食物來源。低 DO 濃度也可能產生厭氧細菌或污染水體的臭味。
水體直接從大氣吸收氧氣,並且由於光合作用從植物吸收氧氣。水保持氧氣的能力受溫度和鹽度的影響。水體主要通過水生植物、動物和微生物的呼吸而消耗氧氣。如果水體較淺,大面積接觸空氣和恆定的運動,未受污染的河流通常包含大量的 DO 供應。如果營養鹽融入過多,大量植物的死亡和營養物質的分解會耗盡氧氣。
水體溶氧
在自然條件下,水體中的氧濃度會波動,但由於人類活動將大量可生物降解的有機物質引入河水中,氧氣會嚴重耗盡。當溪流中的植物產生大量的植物生物量時,營養物質的過量也會使氧氣釋放。
通常,需要氧氣的廢物分解承載被定義為生化需氧量 (BOD) 。 BOD 是通過生物活性來氧化水中有機物質所需的氧氣量的一種度量。因此, BOD 是一個等效指示劑,而不是真正的物理或化學物質。它量測了廢水在溪流中降解時最終需要的總溶解氧濃度。
BOD 也常被分為鈣質組分和含氮組分。這是因為這兩個部分傾向於以不同的速率降解。許多溶解氧的水質模型都需要對多種含碳 BOD(CBODJ 和最終含氮 BOD(NBOD) 或個別氮種濃度進行輸入估算。
需要氧氣分解的污染物可能通過點源排放、非點源排放和地下水溶解排放到河流中。主要點源的 BOD 負載通常是控制和監控的,因此相對容易分析。 BOD 的 非點源承載較難分析。一般來說,從河流流域到河流的任何有機物質的裝載都會導致氧氣消耗需求。有機資料的過量承載可能是由於各種土地利用實踐,再加上暴雨 事件、侵蝕和沖刷造成的。一些農業活動,特別是大規模的動物活動和不當的肥料施用,會導致顯著的生化需氧量荷載。森林栽培和建設中的土地干擾活動,通過對 有機表土的侵蝕,可產生較高的有機承載。最後,都市逕流通常含有來自各種來源的高濃度有機物質
河流中的溶氧
溶氧可能來自大氣或水生植物光合作產生,並且由水中微生物、植物、動物消耗氧。
影響河流中溶氧平衡的主要過程如圖 2.20 所示:
■ 碳質脫氧
■ 含氮脫氧 ( 硝化 )
■ 逸出回歸大氣
■ 沉積物需氧量 �
植物的光合作用和呼吸作用。再氧化是將氧氣引入大多數水域的主要途徑。 氧氣 (O 2 ) 佔大氣的約 21 %並且易於溶解在水中。 DO 中水的飽和濃度是水在給定溫度下可以保持的最大氧含量的量度。當氧氣超過飽和濃度時,它傾向於向大氣中脫氣。當氧氣低於飽和濃度時,它往往會從大氣擴散到水中。根據複數冪功能公式 (APHA1995) ,氧的飽和濃度隨溫度降低。除了溫度,飽和濃度受水鹽度和大氣壓力的影響。隨著水的鹽度增加,飽和濃度降低。隨著大氣壓力的增加,飽和濃度也會增加。大氣與 DO 之間的相互作用由氣相中的分壓梯度和液相中的濃度梯度驅動 (Thomann 和 Mueller1987) 。湍河流和在任一相中混合減少這些梯度和增加的复氧,而在表面上處於靜止,停滯表面或膜減少物理化學特性的复氧。一般而言,天然水中的氧氣轉移取決於以下因素:
■ 由於速度梯度和波動引起的內部混合和湍河流
■ 溫度
■ 風擾動
■ 瀑布,水壩和急河流
■ 表面薄膜
■ 水體深度。
河流復育技術通常利用這些關係,例如通過安置人工級聯以增加再加熱。已經開發了許多用於估計河流再生率係數的經驗公式 ;Bowie 等人提供了詳細的總結。 (1985) 。除了复氧,氧產生插播通過水生植物。通過光合作用,植物從太陽捕獲能量,將二氧化碳固定在有機物質中:
請注意,光合作用也會產生氧氣。植物利用其簡單的光合糖和其他營養物質 ( 特別是氮 [N] ,磷 [P] 和硫 [S] 與少量的常見和微量元素 ) 來操作它們的新陳代謝並建立它們的結構。
大 多數動物的生命取決於植物在光合作用過程中儲存的能量的釋放。在與光合作用相反的反應中,動物消耗植物物質或其他動物並氧化糖,澱粉和蛋白質以促進其新陳 代謝並構建它們自己的結構。該過程稱為呼吸並消耗溶氧。實際的呼吸過程涉及一系列能量轉換氧化還原反應。高等動物和許多微生物依賴於足夠的溶氧作為這些反 應中的末端電子受體,並且在沒有它的情況下不能存活。一些微生物能夠在代謝中使用其他化合物 ( 例如硝酸鹽和硫酸鹽 ) 作為電子受體,並且可以在厭氧 ( 貧氧 ) 環境中存活。
關於河流中 DO 和 BOD 的分析和模擬的詳細信息包含在許多參考文獻中
( 例如, Thomann 和 Mueller1987) ,以及各種經過良好測試的計算機模型。大多數河流水質模型將水體中的 CBOD 與 NBOD( 通常通過氮物種的直接質量平衡表示 ) 和沉積物需氧量或 SOD 分開考慮。 SOD 代表沉積物生物呼吸的需氧量和有機物質的底棲分解。在某些情況下,沉積物和底棲生物對氧的需求可以是河流中總需氧量的很大一部分。在小河流中尤其如此。在低河流量和高溫條件下,效果可能特別嚴重,因為微生物活性趨向於隨著溫度升高而增加。
水體中存在有毒污染物可通過殺死藻類,水生雜草或魚類來間接降低氧氣濃度,從而為耗氧細菌提供豐富的食物。氧耗盡也可能來自不涉及細菌的化學反應。一些污染物引發化學反應,對接收水體施加化學需氧量。

圖 2.20: 由水質模型表示的 BOD 和 DO 的主要動力學過程的相互關係。複雜、相互作用的物理和化學過程有時可以通過模型簡化 , 以便規劃恢復。
2C-2.3 營養成分
除二氧化碳和水外,水生植物 ( 藻類和高等植物 ) 還 需要各種其他元素來支持其身體結構和新陳代謝。與陸生植物一樣,這些元素中最重要的是氮和磷。需要較少量的額外營養素,例如鉀,鐵,硒和二氧化矽,並且通 常不是植物生長的限制因素。當這些化學品有限時,植物生長可能會受到限制。這是河流管理中的一個重要考慮因素。為支持食物鏈,植物生物量 ( 從河流流域產生的河流或從河流中加載 ) 是必需的。然而,藻類和其他水生植物的過度生長會湧現由於植物的呼吸和死亡植物物質的腐爛,非光合作用期間溶氧的消耗會產生不利於水生生物的條件。
淡水系統中的磷存在於顆粒相或溶解相中。兩相均包括有機和無機組分。有機顆粒相包括活的和死的顆粒物質,例如浮游生物和碎屑。無機顆粒磷包括磷沉澱物和吸 附到顆粒上的磷。溶解的有機磷包括由生物排泄的有機磷和膠體磷化合物。可溶性無機磷酸鹽形成 H 2 PO 4 � , HPO 4 2� , and PO 4 3� , , 統稱為可溶性活性磷 (SRP) 是容易獲得的植物,有些稠磷酸鹽形式,例如在洗滌劑中發現的那些,是無機的,但不能直接用於植物吸收。水生植物需要不同量的氮和磷。對於浮游植物,例如,細胞每 pg 葉綠素含有約 0.5 至 2.0 � g 磷,每 � g 葉綠素含有 7 至 10p � g 氮。從這種關係中可以清楚地看出,所需的氮和磷的比例在 5 到 20 的範圍內 ( 取決於各個物種的特徵 ) ,以支持可用營養素的充分利用並使植物生長最大化。當該比例偏離該範圍時,植物不能使用過量存在的營養素。那麼另一種營養就是植物生長的限制性營養素。在經歷過量營養物承載的河流中,資源管理者經常試圖控制限制性營養物的加載量,以防止滋擾狀況。
在水生環境中,氮可以以幾種形式存在 - 溶解的氮氣 (N 2 ) ,氨和銨離子 (NH 3 和 NH 4 + ) ,亞硝酸鹽 (NO 2 - ) ,硝酸鹽 (NO 3 - ) 和有機氮蛋白質物質或溶解或微粒相。就其對水質的直接影響而言,最重要的氮形式是易得的氨離子,亞硝酸鹽和硝酸鹽。因為它們必須轉化為植物更易使用的形式,所以微粒和有機氮在短期內不太重要。
考慮到大氣中含有大約 79 %的氮氣,氮氣可能限制植物生長似乎是不尋常的。然而,只有少數的生命形式 ( 例如,某些細菌和藍 - 綠色藻類 ) 具有與從大氣固定氮氣的能力。大多數植物只能使用氮氣 (NH 3 ,通常作為離子形式銨, NH 4 + 存在於水中 ) 或硝酸鹽 (NO 3 )( 圖 2.21) 。然而,在淡水系統中,水生植物的生長通常受磷的限制而不是氮的限制。出現這種限制是因為磷酸鹽 (PO 4 3 ) 與水中的常見成分形成不溶性配合物 (Ca ++ 和可變量的 OH - , CI - 和 F - ) 。磷也會吸附在粘土和其他沉積物表面的鐵塗層上,因此通過化學過程從水體中除去,導致水體支持植物生長的能力降低。

圖 2.21: 河流生態系統中氮氣的動態和轉化。營養循環從一種形式到另一種形式發生隨著營養投入的變化 , 以及溫度和氧氣的可用量。
整個河流的營養來源
由於人類活動,包括處理後廢水的點源排放和農業和城市發展等非點源,氮和磷都以較高的速率輸送到地表水。在許多發達的河流流域,養分的主要來源是從污水處理廠直接排放處理過的廢物,以及合併下水道溢河流 (CSO S ) 。這種點源排放受到美國污染物排放消除系統 (NPDES) 的管制,通常通過監測得到很好的表徵。 NPDES 要求允許的排放者滿足數字和敘述水質標準在河流中。雖然大多數州沒有營養素的數字標準,但營養素的點源排放被認為是導致河流退化和未能達到敘述水質標準的因素。因此,許多領域對污水處理廠污水 ( 特別是磷 ) 中的養分濃度的限制越來越嚴格。
在許多情況下, NPDES 計畫顯著清理了河流和河流 ; 然 而,即使監管標準越來越嚴格,許多河流仍然不符合水質標準。科學家和監管機構現在明白,許多河流中主要的養分來源是河流河流流域內的非點源,而不是污水處 理廠等點源。導致河流非點污染的典型土地用途是將肥料施用於農田和郊區草坪,不當處理來自牲畜作業的動物糞便,以及在化糞水潭系統中處置人類廢物。來自農 田的暴雨逕流可以以溶解形式和顆粒形式為河流提供養分。
因為它的傾向的吸著到沉澱顆粒和有機物,磷被輸送主要結合侵蝕沉積物地表逕流。無機氮,在另一方面,不吸著強烈和可以運輸在微粒和溶解在地表逕流相。溶解的無機氮也可以通過不飽和區 ( 交河流 ) 和地下水輸送到水體。表 2.6 給出公共點和氮和磷承載的非點源和顯示近似的濃度遞送。請注意,硝酸鹽天然存在於某些土壤中。
河道中的營養循環
由於氮不會很強地吸附沉澱物,因此氮氣很容易在基質和水體之間移動並持續循環。水生生物將溶解的和顆粒狀的無機氮摻入蛋白質物質中。死亡的生物體分解,氮氣以氨離子的形式釋放,然後轉化為亞硝酸鹽和硝酸鹽,再次開始。
磷在淡水環境中經歷連續轉化。一些磷將吸附到水體或基質中的沉澱物中並從循環中除去。 SRP( 通常為正磷酸鹽 ) 被水生植物同化並轉化為有機磷。然後水生植物可能被食蟲和食草動物消耗,而後者又會分泌一些有機磷作為 SRP 。繼續循環, SRP 迅速被水生植物吸收。
表 2.6: 來自共同點和非點源的污染物的來源和濃度。

a. Novotny and Olem (1994). b. 作為有機氮 c. 吸附到空氣中的顆粒物。 d. Omernik (1987). E. 第二次處理
2C-2.4 有毒及有機化學品物質
起毒性的污染物是復育工作的明顯問題。有毒有機化學品 (TOC) 是含有碳的合成化合物,如多氯聯苯 (PCBs) 和大多數農藥和除草劑。許多這些合成的化合物傾向於在環境中持續存在和積累,因為它們在自然生態系統中不容易分解。一些毒性最強的合成有機物,滴滴涕和多氯聯苯,幾十年來一直在美國被禁止使用,但仍然在許多河流的水生生態系統中造成問題。
進入河流的有毒有機化合物
TOC 可通過點源和非點源到達水體。由於允許 NPDES 點源必須滿足水質標準,河道內及全因廢水毒性的要求,在大多數河流持續 TOC 問題都是由於非點加載,存儲在河流和河岸沉積物物資回收,非法傾倒或意外洩漏。有機化學品非點源裝載的兩個重要來源是農業,造林或郊區草坪護理中使用的殺蟲劑和除草劑,以及可能受到污染的城市和工業用地的逕流。
有 機化學品從河流流域地表到水體的運動在很大程度上取決於化學品的特性,如下面縱向視角所述。傾向於污染物吸著強烈土壤顆粒主要運輸與侵蝕泥砂。因此,控制 源區土地利用的泥砂輸送是一種有效的管理策略。具有顯著溶解度的有機化學品可以直接與水流一起運輸,特別是來自不透水的城市表面的暴雨河流。
河流水體中的有毒有機化合物循環
在地球的所有元素中,碳在其與自身形成幾乎無限的穩定共價鍵的能力方面是獨特的:長鏈,分支和環,螺旋螺旋。碳分子可以如此複雜以至於它們能夠編碼用於組織其他碳結構和化學反應調節的信息。
化學工業已經利用它來生產許多有用的有機化學品:塑料,油漆和染料,燃料,殺蟲劑,藥品和其他現代生活用品。這些產品及其相關的廢物和副產品會干擾水生生態系統的健康。了解水生環境中合成有機化合物 (SOC) 的運輸和歸趨繼續挑戰科學家。只有控制這些化學品的行為沿河道的過程的概述呈現在這裡。
碳 - 碳鍵的性質是電子在鍵合原子之間相對均勻地分佈。因此,鏈狀或環狀烴是相當非極性的化合物。
可溶性
這種非極性與水的分子結構不同,水是一種非常極性的溶劑。
根據 � 溶解 � 的一般原則,水中溶解的成分往往是極性的。例如,見證了本章迄今為止討論的幾乎所有無機成分的離子性質。有機化合物如何溶解在水中?有幾種方法。該化合物可以相對較小,因此它最小化了它對水溶液中極性順序的干擾。或者,通過添加極性官能團,化合物可能變得更極性 ( 圖 2.22) 。醇是連接有 -OH 基團的有機化合物 ; 有機酸是具有 -COOH 基團的有機化合物。這些官能團是高度極性的並增加任何有機化合物的溶解度。離子官能團如 COO 在水中的溶解度更高。
提高溶解度的另一種方法是增加芳香性。芳香 � 苯 � 是指環狀化合物 ( 如苯 ) 的離域鍵合結構 ( 圖 2.23) 。 ( 實際上,所有芳香族化合物都可以被認為是苯的衍生物。 ) 因為電子可以自由地在苯分子的 � 環繞 � 上跳動,所以苯及其衍生物更符合水的極性。
一個簡單的例子將說明增強有機化合物水溶性的因素。表 2.7 列出了六種化合物,每種化合物含有六個碳。己烷是一種簡單的烴,一種烷烴,其溶解度為 10mg/L. 簡單地通過添加單個 -OH 基團,其將己烷轉化為醇己醇,溶解度增加至 5,900mg/L. 您可以將己烷彎曲成環狀烷烴結構,稱為環己烷。形成環形成環己烷比己烷小,增加其溶解度,但僅為 55 毫克 / 升。通過形成六碳苯分子使環形芳香化,一直增加溶解度至 1,780mg/L. 向苯中加入 -OH 形成苯酚導致溶解度的另一個顯著增加 ( 至 82,000mg/L) 。向苯環中加入氯原子會降低其芳香性 ( 氯化物抑制跳舞電子 ) ,因此氯苯 (448mg/L) 的溶解度小於苯。
圖 2.22: 不同功能組的相對水溶性。水中污染物的溶解性在很大程度上決定了污染物對水質的影響程度。


圖 2.23 :芳香烴。 苯由於其�芳族�結構而可溶於水。
吸附
在 20 世紀 40 年代,一個年輕的製藥行業尋求開發可以在消化液和血液中運輸的藥物 ( 兩者都基本上是水溶液 ) ,也可以擴散穿過細胞膜 ( 部分地具有相當非極性的特徵 ) 。該行業開發了一個參數來量化潛在藥物的極性與非極性特徵,他們將該參數稱為辛醇 - 水分配係數。基本上,他們將水和辛醇 ( 八碳醇 ) 放入容器中,加入感興趣的有機化合物,並將組合物震動。休息一段時間後,將水和 OCtanol 分開 ( 兩者都不是在其他非常可溶的 ) ,和有機化合物的濃度可在每個階段中被測量。所述辛醇 - 水分配係數, K ow ,被簡單地定義為:
K ow= 辛醇濃度 / 水中濃度
水溶性和 Kow 之間的關係如圖 2.24 所示。通常我們看到像滴滴涕和多氯聯苯非常不溶的化合物具有非常高的值 ķ 河流。或者,有機酸和小有機溶劑如 TCE 是相對可溶的並且具有低 K ow 值。
K ow 已經確定了許多化合物的辛醇 - 水分配係數,可用於了解水和生物群之間以及水和沉積物之間 SOC 的分佈。具有高 Kow 的化合物傾向於積聚在魚組織中 ( 圖 2.25) 。所述沉積物 - 水分配係數,通常表示為 K d ,是在沉積物 - 水混合物在平衡時的濃度的沉積物在水中的濃度的比率來定義:
K d = 沉積物濃度 / 水中濃度
有人可能會問,對於給定的 SOC ,該係數是否恆定。圖 2.26 顯示了兩種土壤中兩種多芳烴的 Kd 值。
對於 pyrene( 由四個結合在一起的苯環組成 ) , Kd 比在 300 到 1500 之間變化。對於菲 ( 由三個被結合在一起的苯環組成 ) , Kd 從大約 10 到 300 之間變化。顯然, Kd 不是常數兩種化合物的值。但是, Kd 的確與各種沉積物中有機碳的含量有關。看起來恆定的不是 Kd 本身,而是 Kd 與沉積物中有機碳比例的比率。該比率稱為 Koc :
Koc = K d / 有機碳含量
沉積物中的各種功已將 Koc 與 Kow 以及水溶性相關 ( 表 2.8) 。使用 Kow , Koc 和 Kd 描述水之間 SOC 的分配,沉積物已顯示出一定的效用,但這種方法不適用於所有系統中所有有機分子的吸附。某些 SOC 的吸附通過氫鍵發生,例如發生在陽離子交換或金屬對沉積物的吸附中 ( 圖 2.27) 。吸附並不總是可逆的。或至少在發生吸附後,解吸可能會非常緩慢。
表 2.7: 六碳化合物的溶解度。


圖 2.24: 八醇 /H 2 O 分區係數與水溶性之間的關係。
水中的相對溶解性是物質的 " 水分係數 " 。
圖 2.25: 八醇 / 水隔 (P 八度 ) 系數與鮭魚肌肉生物吸收因數 (BCF) 之間的關係。水質可以通過魚組織中污染物的積累來推斷。
![2-25%C2D%B3%BD%A6%D9%A6%D7%A5%CD%AA%AB%A7l%A6%AC%A6]%BC%C6](S02-河流地貌過程與功能.files/image075.jpg)
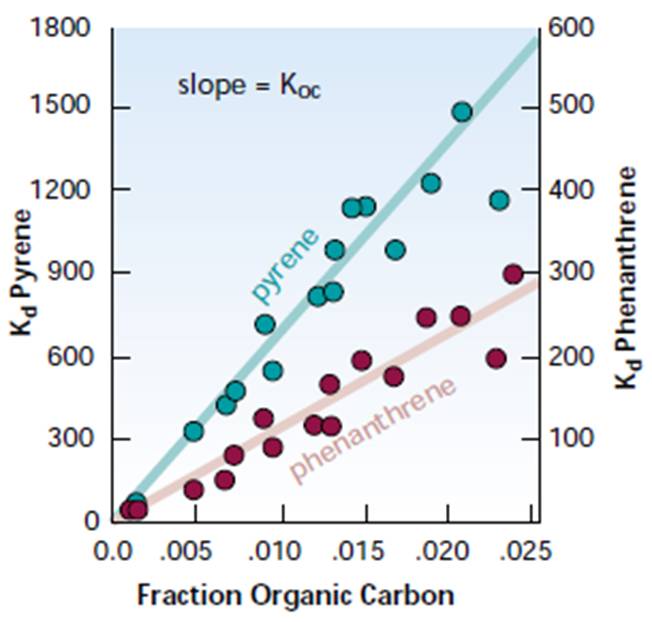
圖 2.26: 苯甲酸酯、苯丙烯和餾分有機碳之間的關係。沉積物與水的污染物濃度 (Kdd) 與可用有機碳的含量有關。
揮發
有機化合物通過揮發過程從水分配到空氣中。空氣 - 水分配係數,亨利定律常數 (H) ,定義為平衡空氣中 SOC 濃度與其在水中濃度的比值:
H= 空氣中的 SOC 濃度 / 水中的 SOC 濃度。
� SOC � = 合成有機化合物 SOC 的亨利法則常數可以根據化合物的蒸氣壓與其水溶性的比率來估算。具有固有揮發性的有機化合物 ( 通常是低分子量溶劑 ) 具有非常高的亨利定律常數。但即使是蒸氣壓很低的化合物也可以分解到大氣中。例如,滴滴涕和多氯聯苯具有適度的亨利定律常數,因為它們在水中的溶解度很低。這些 SOC 也具有高 ķ d 值,因此可以被來在用微粒物關聯空中。

圖 2.27: 涉及天然有機物和礦物表面的兩種重要氫結合。有些污染物是由沉澱物顆粒的 , 這些沉積物顆粒通過化學粘結吸附到其表面。

圖 2.28: 電磁輻射能量與一些選定的鍵式能量相比。光透過光解打破某些化合物的化學鍵。
降解
SOC 可以轉化為各種降解產物。這些降解產物本身可能會降解。最終的降解或礦化導致有機碳氧化成二氧化碳。主要的轉化過程包括光解,水解和氧化還原反應。後者通常由生物系統介導。
光解是指通過光能破壞化合物。光的能量與其波長成反比 ( 圖 2,28) 。長波光缺乏足夠的能量來破壞化學鍵。短波光 (X 射線和伽馬射線 ) 非常具有破壞性 ; 幸運的是,對於地球上的生命,這種類型的輻射很大程度上被我們的高層大氣所消除。可見光譜附近的光線到達地球表面,可以破壞 SOC 中常見的許多鍵。揮發後有機溶劑的命運通常是在地球大氣中的光解作用。光解對於河流水中 SOC 的降解也很重要。
水解是指用水分解有機分子。基本上水進入分子的極性位置並插入自身, H+ 進入母體分子的一部分, OH- 進入另一部分。然後將這兩部分分開。一組稱為酯的 SOC 特別容易被水解降解。許多酯已經作為殺蟲劑或增塑劑生產。
氧化還原反應是促進生物圈中大部分新陳代謝的因素。 SOC 通常被認為是減少碳的來源。在這種情況下,什麼是需要對降解是一種代謝系統與 APPR 需用合適的化合物的氧化 酶 。還需要足夠的其他營養素和末端電子受體。
微生物無謬誤的原則非正式地提到了這樣一種觀點,即在提供潛在食物的情況下,微生物群落將發展代謝能力以將該食物用於生化能量。然而,並非所有降解反應都涉及 SOC 的氧化。一些最有問題的有機污染物是氯化化合物。
氯化 SOC 不 是天然存在的,因此微生物系統通常不適合它們的降解。氯是一種極具電負性的元素。氯的電負性指的是它吸收電子的傾向。這種趨勢解釋了為什麼氯作為陰離子存 在以及為什麼附著的氯化物會降低芳環的溶解度。鑑於這種特性,生物系統難以氧化氯化化合物。因此,降解的第一步通常是還原脫氯。通過還原化合物 ( 即通過給予電子 ) 除去氯。除去氯後,降解可以沿著氧化途徑進行。因此,氯化 SOC 的降解可能需要一系列還原和氧化環境,水可能在河流和潛河流區之間移動時經歷。
SOC 的整體退化通常遵循複雜的途徑。圖 2.29 顯示了單一母體農藥的代謝反應的複雜網絡。水解,還原和氧化都涉及 SOC 的降解,並且降解產物的分佈和行為在空間和時間上可以是極其可變的。
化學後果很少是大多數復育行動的直接目標。改變化學過程和屬性的計畫通常側重於改變對復育目標至關重要的物理和生物特徵。
2C-3 重金屬的毒性濃度
已 經建立了各種天然存在的金屬,從砷到鋅,當以足夠的濃度存在時對各種形式的水生生物有毒。大多數金屬的水體毒性的主要機制是在鰓表面吸附。雖然一些研究表 明,顆粒金屬可能會導致毒性,可能是因為諸如鰓表面解吸等因素,溶解的金屬濃度最接近水生物可利用的金屬部分。因此,目前的 EPA 政策是應該使用溶解的金屬濃度設定和測量水質標準的符合性 (40CFR22228-22236 , 1995 年 5 月 4 日 ) 。對於大多數金屬,溶解的部分相當於無機離子部分。對於某些金屬,特別是汞,溶解的部分也可能包括與有機粘合劑複合的金屬 ( 例如,甲基汞,可以在產甲烷細菌的沉積物中產生,是可溶的和劇毒的,並且可以通過食物鏈積累 ) 。
整個河流的重金屬的毒性濃度
與合成有機化合物不同,有毒金屬是天然存在的。與合成有機物一樣,金屬可以從點和非點源加載到水體中。污染物,如銅,鋅和鉛通常在廢水處理廠河流出物的關注,但都是 Required 的 NPDES 方案,以滿足下數字的水質標準。許多有毒金屬在大多數土壤中以顯著濃度存在,但是以吸附的非生物可利用形式存在。沉積物通常會將大量金屬 ( 如鋅 ) 引入水體中。它是那麼是否事河道內條件促進生物利用溶解形式的金屬。
非點源金屬首先反映了河流流域土壤的特徵。此外,許多較老的工業區的某些金屬的土壤濃度由於過去的工業實踐而升高。金屬從土壤到河流流域的運動在很大程度上取決於沉積物的侵蝕和輸送。
在某些河流流域,酸性礦井排水提供了主要的金屬承載來源。高酸度增加了許多金屬的溶解度,礦物往往富含礦物質。因此,廢棄的地雷是許多河流中有毒金屬承載的持續來源。
河流水體中的重金屬的毒性濃度
大多數金屬具有一傾向離開溶解相並附著於懸浮的顆粒物質或形成不溶性的沉澱物。將金屬分成顆粒形式的條件 ( 懸浮沉積物,溶解和顆粒狀有機碳,碳酸鹽,碳酸氫鹽和其他複雜金屬離子的存在 ) 降低了金屬的潛在生物利用度。此外,鈣可以減少金屬吸收,顯然是通過與金屬競爭鰓膜上的活性攝取部位。 pH 也是金屬生物利用度的重要水質因素。通常,金屬溶解度在接近 neutralpH 值時比在酸性或高鹼性水中低。
表 2.8: 各種污染物沉積物吸附係數 (K OC 的回歸方程式。

a.Koc = 土壤 ( 或沉積物 ) 的吸附係數; S = 水溶性; Kow = 辛醇 - 水分配係數; BCF(f)= 流水測試中的生物濃縮係數; BCF(t)= 來自模型生態系統的生物濃縮因子; P = 降落傘; N = 分子中可參與氫鍵形成的位點數。
b. 編號 = 用於獲得回歸方程的化學藥品數量。
c. r2 = 回歸方程的相關係數。
d. 最初根據 Kom 給出的方程式。 關係 Kom = Koc / 1.724 用於根據 Koc 重寫方程。
e. 不可用。 f 未指定用於獲得回歸方程的特定化學品。
圖 2.29: 單親農藥的代謝反應。粒子分解通過水解、氧化、還原和光解的過程。

2C-4 土壤的生態功能
土壤是一種生活和動態的資源,支持生命。它由不同大小的無機礦物顆粒 ( 粘土,淤泥和沙子 ) ,不同分解階段的有機物,多種生物,各種水溶性離子以及各種氣體和水組成。這些組件各自具有其自身的物理和化學特性,可以支持限制特定的生命形式。
土壤可以是礦物質或有機物,這取決於哪種物質佔土壤基質中的較大百分比。礦質土壤在岩石風化的物質中生長,而有機土壤則在腐朽的植被中生長。
兩種土壤通常形成大致平行於土壤表面的視野或層。土壤可以產生的特定生態位或條件的極端多樣性使得各種動植物群落在這些條件下進化和生存。
土 壤,特別是河岸和濕地土壤,包含和支持土壤表面上下的非常多樣的動植物。在土壤表面下方可以找到各種各樣的特殊生物,比地上數量多出幾個數量級。通常,在 地上看到的生物是更高形式的生命,例如植物和野生動物。然而,在地上和地下,絕大多數生命由植物根系組成,負責支撐植物的地上游分 ; 許多以死有機物為生的昆蟲,軟體動物和真菌 ; 和無數的細菌,可以生活在土壤中發現的各種能源。
重 要的是識別土壤邊界並理解在河道內發生的土壤性質和功能的差異,以便識別復育的機會和限制。洪氾區和露台土壤通常是種群數密集和農業發展密集的地區,因為 它們平坦的斜坡,靠近水和自然肥力。在發達地區規劃河道河流復育計畫時,重要的是要認識到這些變化並考慮它們對目標的影響。
土 壤在整個景觀中發揮重要作用。土壤最重要的功能之一是為生物提供物理,化學和生物環境。土壤支持植物和動物生產力的生物活動和多樣性。土壤還調節和分配水 的河流動以及景觀中養分和其他元素的儲存和循環。它們過濾,緩衝,降解,固定和解毒有機和無機物質,並提供生物體所需的機械支持。這些水文,地貌和生物功 能涉及幫 助建立和維持河道的過程。
2C-4.1 土壤微生物學
有機物質是土壤微生物的主要能量來源。土壤有機質通常佔礦物表土總重量的 1 %至 5 %。 它由原始組織,部分腐爛的組織和腐殖質組成。土壤生物消耗能量和構建組織的根和營養碎屑。當原始有機物被微生物分解和改性時,形成凝膠狀,更耐藥的化合 物。這種物質叫做腐殖質。它通常為黑色或棕色,並以膠體形式存在,一組懸浮在凝膠中的小的不溶性顆粒。少量的腐殖質大大增加了土壤保持水分和養分離子的能 力增強植物生產。腐殖質是土壤中大量可行的微生物種群的指標,它增加了可用於營養復育的選擇。
細菌在支持植物生長的有機交易中發揮著至關重要的作用。它們負責三種必要的轉化:反硝化,硫氧化和固氮。微生物將硝酸鹽還原為亞硝酸鹽,然後還原為氣態氮,稱為反硝化。含水量為 60 %通常會限制反硝化作用,該過程僅發生在 5 � C 至 75 � C 的土壤溫度下。優化反硝化速率的其他土壤性質包括 pH 值在 6 之間 8 ,土壤通氣低於土壤中生物的生物需氧量,足量的水溶性碳化合物,土壤中容易獲得的硝酸鹽,以及開始反應所需的 酶 的存在。
2C-4.2 景觀和地形位置
土壤性質隨地形位置而變化。海拔差異通常標誌著河道中土壤和排水條件的界限。不同的地貌通常具有不同類型的沉積物。地表和地下排水模式也隨地形而變化。
■ 活動河道的土壤。活動河道形成河流河道中最低且通常最年輕的表面。在這些表面上通常沒有形成土壤,因為形成河底和堤岸的未固結物質經常被侵蝕,運輸和再沉積。
■ 活躍洪氾區的土壤。河道中的下一個最高表面是活動洪氾區的平坦沉積表面。這個表面經常在 3 年內每 2 年發生洪水,因此會接收沉積物沉積。
■ 岸側自然堤壩的土壤。在洪水期間,通過沉積較大的懸浮沉積物從堤岸河流中沉積,在河流附近建造岸側自然堤壩。溫柔的背影 -
■ 坡面發生在岸側自然堤壩的洪氾區一側,因此洪氾區在遠離河流的地方變得最低。由於污水區域的沉積物輸送能力下降,母體物質的粒度遠離河流。
■ 地形氾濫平原的土壤。活動洪氾區內外的略高的區域被定義為地形洪氾區。它們通常比活躍的洪氾區更少地被淹沒,因此土壤可能比活躍的洪氾區上的年輕土壤表現出更多的剖面。
■ 梯田的土壤。被遺棄的洪氾平原或梯田是河道中的下一個最高表面。這些表面很少氾濫。一般來說,露台土壤比河灘地土壤更粗糙,更自由地排水,並且與河流過程分開。
經 過仔細檢查,洪氾平原礦床可以揭示特定河流流域的歷史事件。土壤剖面開發提供了一個場地的近期和地質歷史的線索。複雜和複雜的分析方法,如碳定年,花粉分 析,某些同位素的比例等,可用於拼湊一個地區的歷史。侵蝕或沉積的循環有時可能與森林火災或高降雨或低降雨期等災難性事件有關。文明的歷史影響,例如廣泛 的農業或剝奪森林覆蓋,有時也會在土壤中留下可識別的證據。
2C-4.3 土壤溫度和濕度關係
土壤溫度和水分控制土壤中發生的生物過程。在考慮河流復育計畫的目標時,平均和預期的降雨和極端溫度是關鍵的信息。年平均土壤溫度通常與年平均氣溫非常相 似。土壤溫度確實經歷由太陽輻射,天氣模式和氣候引起的日常,季節和年度波動。土壤溫度也受到方位,緯度和海拔的影響。
土壤濕度條件季節性變化。如果將植被物種和組成的變化視為河流復育計畫的一部分,則應構建一個比較月降雨量和植被蒸發量的圖表。如果地下水位和毛細管邊緣 低於預測的生根深度,並且圖表顯示可用水的不足,則可能需要灌溉。如果沒有補充水,則必須考慮不同的植物種類。
土壤水分梯度可以從 100 %的水體減小到高旱土地的近似 0% ,河岸帶演替梯度從從河流底部,穿過河岸帶,並進入相鄰的內陸高地,土壤水分的漸變差異,使植被呈現多樣化的層次。土壤水分直接影響河岸帶、過渡帶和高地帶的生態群落的特徵。這些生態差異導致沿著河道存在兩條生態交錯帶,水生濕地 → 河岸生態交錯帶 (eco-tone) 和非濕地河岸 / 洪氾區生態交錯帶。其中增加河岸帶的邊緣效應,從而增加該地區的生物多樣性。
2C-4.4 濕地土壤
潮濕或 � 含氫 � 的 土壤對植物生命提出了特殊的挑戰。濕地土壤中存在含水土壤,在高地上發現的大多數物種無法生存的物理和化學條件發生如此劇烈的變化。因此,濕地中的動植物 群落構成了巨大的變化。不同的和獨特的,尤其是在受到永久或長期飽和或浸水濕地。含氫土壤被定義為那些飽和,水淹或積水足夠長的時間,在生長季節,開發厭 氧條件中上游。這些厭氧條件影響植物的繁殖,生長和存活。氫的形成背後的驅動過程在生長季節,土壤在地表附近長時間 ( 通常超過 7 天 ) 充滿水分和 / 或土壤飽和度。
以下主要關注礦物質水合土壤性質,但河道中可能存在有機土壤,如泥炭和淤泥。
在 充氣土壤環境中,大氣氧氣通過氣體擴散進入表層土壤,因為土壤孔隙大部分充滿空氣。在充分排水的高地中發現了充氣土壤,並且通常所有區域都具有遠低於根區 的地下水位。在飽和土壤中,孔隙充滿水,與大氣相比,氣體非常緩慢地擴散氣體。只有少量的氧氣可以溶解在土壤水分中,然後分散到土壤的頂部幾公分處。在這 裡,土壤微生物迅速耗盡了將有機殘留物氧化成二氧化碳的所有可用氧。該反應產生的厭氧化學還原,其中氧化的化合物變更為可溶於還原的化合物環境,並且還對 許多植物有毒。擴散速度很慢,在這種情況下不能重新建立氧化條件。在浸水厭氧環境中涉及有機物分解的類似微生物反應產生乙烯氣體,其對植物根部具有高毒性並且比缺氧具有更強的效果。在使用所有的游離氧之後,厭氧微生物減少了土壤的其他化學成分,包括硝酸鹽,氧化錳和氧化鐵,從而在土壤中進一步減少了條件。
延長的厭氧還原條件導致形成易於看到的還原跡象。在潮濕的土壤中遇到的典型的灰顏色是還原鐵的結果,並且是公知的作為潛育土壤。在鐵氧化物耗儘後,硫酸鹽 被還原成硫化物,產生濕土壤的臭雞蛋氣味。在極度淹水的條件下,二氧化碳可以減少為甲烷。甲烷氣體,也稱作為 � 沼澤氣 � 可在夜間看到的,因為它發出螢光。
一些濕地植物已經進化出特殊的機制來補償它們的根浸入缺氧環境中。例如,睡蓮通過在一天的高溫期間關閉它們的氣孔來迫使整個植物內的氣體交換,以提高特殊導電組織 ( 通氣組織 ) 內的氣壓。這個過程傾向於將大氣氧氣引入根冠深處,使重要組織保持活力。大多數暫時性濕地植物只是將其根系保持在土壤表面附近,以避免深層地區的厭氧條件。例如,莎草就是如此。蘆葦與香蒲可提供大量的氧氣進入底泥,同時在洪水淹到這地區時能夠有快速的復原性。
當 土壤持續飽和時,反應可以在整個土壤剖面中均等地發生,而不是在水位波動的濕土壤中。這會產生幾乎沒有分區的土壤,物質往往更均勻。深度遇到的紋理的大多 數差異與通過河流動水沉積期間按尺寸分類的沉積物的分層有關。粘土的形成傾向於在原位發生,並且在型材內發生很少的易位,因為基本上沒有水通過土壤移動以 輸送顆粒。由於濕土的反應性,粘土的形成往往比高地快得多。
土壤,其被季節性飽和的或具有不同的在一個波動水表結果水平輪廓內。由於水經常通過輪廓排出,它使顆粒易位並將可溶性離子從一層傳遞到另一層,或完全從輪 廓中傳出。通常,這些土壤在表面附近有一層厚的地平線,剝去了包括鐵在內的所有可溶性物質 ; 稱 為耗盡矩陣。季節性飽和的土壤通常在表面積累大量有機物質,顏色幾乎為黑色。有機物添加到陽離子中土壤的交換能力,但由於剝離和氫離子過多,鹼飽和度很 低。在非飽和時期,有機物質暴露於大氣氧,並且可以發生需氧分解,這導致氫離子的大量釋放。季節性潮濕的土壤也不能很好地保留基礎金屬,並且在乾燥期後可 以在濕循環中釋放高濃度的金屬。
潮濕的土壤指標往往停留在長期的土壤剖面的時間段 ( 甚至引河流後 ) ,揭示其盛行的歷史條件。這些指示劑的實例是銹色鐵沉積物,其一度被還原形式的水轉移。由過去的河流沉積循環或由濕地情況引起的剝離土壤的區域的有機碳分佈是非常長壽的特徵。
2C-5 摘要
本節僅簡要概述了各種複雜的化學反應 ; 儘管如此,復育我們應該明白兩個關鍵點:
■ 如果生態系統存在水質限制,復育物理棲息地無法復育系統的生物完整性。
■ 復育活動可能以各種複雜的方式相互作用,水質影響水質壓力源的傳遞和影響。
表 2.9: 選定溪流恢復和河流流域管理做法的潛在水質影響。
表 2.10 :土壤中常見的生物類群。


2D 生物群落特徵
成功的河流復育基於對不同時間尺度的物理,化學和生物過程之間關係的理解。通常,人類活動加速了這些過程的時間進展,導致不穩定的河流動模式和改變的河道的生物結構和功能。本節討論了與地貌,水文和水質 過程相關的河道的生物結構和功能。還討論了河流流域與河流之間的相互關係,以及擾亂這些相互關係的原因和影響。第 7 章提供了評估河道功能的指標和方法。
2D-1 陸域生態系
河道的生物群落由陸地和水生生態系統的特徵決定。因此,對河道中生物群落的討論始於對陸地生態系統的審查。
2D-1.1 土壤的生態作用
陸地生態系統從根本上與土壤中的過程聯繫在一起。土壤儲存和循環養分及其他元素的能力取決於土壤的性質和微氣候 ( 即水分和溫度 ) 以及土壤的生物群落 ( 表 2.10) 。這些因素也決定了其在過濾、緩衝、降解、固定和解毒其他有機和無機物質方面的有效性。
2D-1.2 陸域植被
河道生態系統的生態完整性與構成和圍繞廊道的植物群落的完整性和生態特徵直接相關。這些植物群落是生物群落的寶貴能源,為周圍的水生和陸地生態系統提供物 理棲息地和適度的太陽能通量。在充足的水分,光照和溫度下,營養群落以活躍的生長 / 生 產,衰老和相對休眠的年度週期生長。增長期受到補貼通過偶然的太陽輻射,驅動光合作用過程,無機碳通過該過程轉化為有機植物物質。這種有機物質的一部分作 為地上和地下生物質儲存,而有機物質的大部分每年通過衰老,分餾和以葉子,樹枝和腐爛根的形式浸出到有機土壤層中而損失。。這種有機部分富含微生物菌群和 微型動物的生物活性,是可利用的碳,氮,磷和其他營養物質的主要儲存和循環水潭。
營養群落的分佈和特徵取決於氣候,水的供應,地形特徵以及土壤的化學和物理特性,包括水分和養分含量。植物群落的特徵直接影響著動物群落的多樣性和完整性。覆蓋面積大且垂直和水平結構特徵多樣的植物群落可以支持比相對同質的植物群落 ( 如草地 ) 更多樣化的動物群落。結果是在花卉和動物群落之間存在複雜的時空關係,這些群落的河流生態特徵反映了近期歷史 (100 年或更短 ) 的 景觀物理條件。陸地植被的數量及其物種組成可直接影響河道特徵。河床中的根係可以結合堤岸沉積物和適度的侵蝕過程。落入河流中的樹木和較小的木質碎片會使 河流動偏轉並在某些點引起侵蝕並在其他點沉積。因此,木質碎屑堆積可以影響水潭分佈,有機物質和營養物保留,以及作為重要魚類和無脊椎動物水生群落的微棲 地的形成。水流也可能受到陸地植被豐度和分佈的影響。除去植被的短期影響可能導致當地地下水位短期上升,原因是蒸發蒸騰減少和進入河流的額外水量。然而, 從長期來看,在去除植被後,河流的基河流會減少,水溫會上升,特別是在低階河流中。此外,植被的移除可能導致土壤溫度和結構的變化,導致運動減少水進入並 通過土壤剖面。表面凋落物的減少和土壤中有機物質的逐漸減少也會增加地表逕流和減少滲透。在大多數情況下,最明顯的植被功能是影響魚類和野生動物的功能。 在景觀層面,本土覆蓋類型的碎片化已經顯示出對野生動物的顯著影響,通常偏向於機會主義物種而不是需要大塊連續棲息地的物種。在 some 系 統中,相對而言廊道連續性的小斷裂可能對動物運動或河流條件適合支持某些水生物種產生重大影響。在其他情況下,建立與本機系統結構不同或者配置不當的廊道 可能同樣具有破壞性。狹窄的廊道基本上是邊緣棲息地可能會鼓勵通才物種,巢寄生蟲和捕食者,並且,在建立跨越動物運動的歷史障礙的廊道時,它們可以破壞區 域動物群落的完整性 (Knopf 等人, 1988) 。
河流流域景觀尺度
河 流流域植物群落的生態特徵和分佈影響水,沉積物,養分和野生動物的遷移。河道提供與景觀的其他功能的鏈接。鏈接可能涉及源頭和谷底生態系統之間的連續廊道 或地面系統之間的周期性相互作用。野生動物使用廊道驅散幼魚,遷移,並在其家鄉的部分區域之間移動。天然來源的廊道是首選,包括河流和河流、河岸帶、出山 口,峽谷和狹窄的河道 (Payne 和 Biyant1995) 。
了解河流 - 河岸生態系統與河流 - 河灘地生態系統之間的差異非常重要。河流 - 河岸生態系統中的洪水是短暫且不可預測的。河岸帶為河道提供養分、水和沉積物,河岸植被調節溫度和光照。在河流 - 洪氾區生態系統中,洪水往往更具可預測性和持久性,河道是水,沉積物和無機養分的供給者。
洪氾區以及渾濁和較冷的河道水的河流入會影響淹沒的洪氾平原的光穿透和溫度。
河道景觀寬度
在河道尺度上,植被的組成和再生模式以水平複雜性為特徵。沿無約束河道的洪氾區通常植被茂密的植物群落,其組成隨可用的地表水和地下水而變化,洪水、火災和主要風的差異模式,沉積物沉積以及建立植被的機會。
美國南部,中西部或東部的廣闊洪氾區可以在複雜的鑲嵌塊中支持數十個相對不同的森林群落,反映出土壤類型和洪水特徵 ( 例如,頻率,深度和持續時間 ) 的 細微差別。相反,雖然某些西部河流水系可能僅支持少數木本物種,但由於河流對基質的不斷重新加工,這些系統可能在結構上複雜,這產生了不同年齡的林分的鑲 嵌塊。可以查看側河道,牛軛湖和其他地形變化的存在作為河道層面結構變化的要素。受限河流河道沿岸的河岸區域可能主要由高度植被組成,這些植被由與河流特 徵無關的過程組織,但這些區域可能對河流生態系統產生相當大的影響。
如第 1 章所述,河流連續體概念通常也適用於河岸廊道的植被組成部分。河岸植被表現出跨越式的梯度 ( 橫跨谷 ) 和河岸內 ( 縱向,高度 ) 梯度 (Johnson 和 Lowe1985) 。在西部,河道植被的生長因 � 涼風效應 � 而增加,當涼爽潮濕的空氣從高海拔地區溢出下坡時 ( 圖 2.30) 。這種較涼爽的空氣在峽谷中沉澱,並產生比周圍斜坡更濕潤的微棲地。這些峽谷也可作為水道。濕潤,涼爽的組合土壤和大氣條件有利於低於正常海拔的植物和動物物種,通常在分離種群或不會發生的地區。
植物群落
動物群落對植物特性的敏感性已得到公認。許多動物物種與特定植物群落相關,許多動物物種需要這些群落的特定發育階段 ( 例如,老齡生長 ) ,並且一些動物物種依賴於這些群落內的特定棲息地元素 ( 例如,障礙物 ) 。河濱植物群落的結構也直接通過提供適當的有機物質投入到水生食物網,通過遮蔽水表面和沿著水岸提供蓋,並通過影響影響的水生生物的棲息地,可以從內部複雜性來看待植物群落 ( 圖 2.31) 。複雜性可包括植被層數和構成每層的物種 ; 物種之間的競爭互動 ; 和碎屑成分的存在,如墊料,倒塌的木材和障礙物。植被可以含有大喬木、小喬木、灌木、爬藤類、草類等、苔蘚等層。微觀圖像浮雕和水對當地水窪的能力也可被視為特徵性結構組件。
生物學文獻中地層多樣性或樹葉高度多樣性概念中描述的垂直複雜性對於 Carothers 等人對鳥類棲息地的研究非常重要。 (1974 年 ) 沿著佛得角河,在河流中游的五級序、六級序河流。研究結果表明,河岸鳥類物種多樣性與河岸植被的葉片高度多樣性之間存在高度相關性 (Carothers 等, 1974) 。 Short(1985) 證明,更多結構多樣的營養棲地支持更多的行會 guild( 同功群 )( 社區中具有密切相關生態位的物種群 ) ,因此物種數量更多。
植 被結構的種類和年齡組成也非常重要。簡單的植物結構,例如沒有木質過度的草本層或沒有較小尺寸等級的老木質河岸樹木,為覓食同功群創造了更少的利基。同功 群越少,物種越少。植被的質量和活力可以影響果實、種子,嫩芽,根和其他營養物質的生產力,這些物質為野生動物提供食物。較差的活力可導致較少的食物和較 少的消費者 ( 野生動植物 ) 。
增加河邊植被類型的斑塊大小 ( 面積 ) ,增加木質河岸樹大小類別的數量,增加原生河岸依賴植被的物種和生長形式 ( 草本植物,灌木,樹木 ) 的數量可以增加行會 guild( 同功群 ) 和牧草的數量,導致物種豐富度和生物量 ( 數量 ) 增加。復育技術可以改變上述因素。
河流廊道內水平複雜性對某些動物物種的重要性也已得到很好的證實。例如,南部河灘地森林的特徵組成,結構和地形複雜性提供了許多越冬水禽在適當的時間滿足其生命週期特定要求所需的資源和覓食條件 (Fredrickson1978); 據報導,洪氾平原棲息地的其他脊椎動物和無脊椎動物也存在類似的複雜關係 (Wharton 等, 1982) 。在乾旱的西部的部分地區,河岸系統中獨特的植被結構可以發揮作用與周圍的高地一起,為許多動物提供必要的棲息地 (Knopf 等, 1988) 。即使在組成簡單的河岸系統中,不同的開發階段也可以提供不同的資源。
植物群落分佈在洪氾平原上,涉及洪水深度,持續時間和頻率,以及土壤和排水條件的變化。一些植物品種,如棉白楊、柳樹和銀槭,適合於新沉積的沉積物的定殖,並且可能需要 veiy 期間的短暫時期特定洪水衰退類型成功建立種子附著 (Morris 等, 1978 , Rood 和 Mahoney , 1990) 。得到的植被分布是在河流的活動曲折帶內以不同的間隔和位置建立的均勻樹齡林中的一個。其他物種,如禿柏,尤其與河流切斷河道段時形成的牛軛湖相關,而其他物種則與洪氾平原內的微地形變化有關,反映了河道穿過河道的緩慢遷移的景觀。
植 物群落是動態的,並隨著時間而變化。特定植被類型的不同再生策略導致在干擾之後植物演替的特徵模式,其中適應裸土和豐富光的先鋒物種逐漸被可以在更多陰影 和受保護條件下再生的更長壽命物種所取代。新的干擾重置了演替過程。在河道內,洪水,河道遷移以及在某些生物群落中,火災通常是干擾的主要自然來源。復育 我們應該了解河流中的自然繼承模式
通過種植耐寒的早期演替物種來穩定侵蝕的河床,同時規劃最終用更長壽和更高演替的物種替代這些物種,應利用演替過程。

圖 2.30 :峽谷效應。涼爽潮濕的空氣沉澱在峽谷中,在周圍的山坡上形成了小棲地。
2D-1.3 陸域動物群落
野生動物使用河流廊道比任何其他棲息地類型更為頻繁 (Thomas 等人 1979) ,並且是野生動物種群,特別是大型哺乳動物的主要水源與食物來源。例如,亞利桑那州 60 %的野生動物物種依賴於河岸地區生存 (Ohmart 和 Anderson1986) 。在猶他州和內華達州的大盆地地區, 363 個已確定的陸地脊椎動物物種中有 288 個依賴於河岸帶 (Thomas 等, 1979) 。由於它們對高地和河岸物種的廣泛適應性,與草原草原相關的中西部河道支持比相關高地更多樣化的野生動物。河道在維護所有脊椎動物群體的生物多樣性方面發揮著重要作用。
河道的動物群構成是食物、水、覆蓋和空間佈局相互作用的功能 (Thomas 等, 1979) 。這些棲息地組件以多種方式相互作用,以提供河道的八個棲息地特徵:
■ 存在永久性水源。
■ 高初級生產力和生物量。
■ 覆蓋類型和食物供應的戲劇性時空對比。
■ 關鍵的微氣候。
■ 水平和垂直棲息地多樣性
■ 最大化的邊緣效果。
■ 有效的季節性遷移路線。
■ 植被斑塊之間的高度連通性。
河道為許多形式的野生動物提供了最佳棲息地,因為靠近水源和生態社區,該社區主要由該國許多地區的大喬木組成,提供食物來源,如花蜜,柳絮,芽,水果和種子 (Harris1984) 。上游水源,養分和能源最終有利於下游地區。反過來,魚類和野生動物在其移動和遷徙過程中返回並將一些營養和能量分散到高地和濕地 (Harris1984) 。
水 對於草原地區的動物群尤為重要,其中河道是景觀中唯一天然存在的永久性水源。這些相對潮濕的環境有助於河岸地區的高初級生產力和生物量,這與周圍的覆蓋類 型和食物來源形成鮮明對比。在這些區域,河道提供了關鍵的微氣候,通過提供水,陰影,蒸發蒸騰和覆蓋來改善高地的溫度和水分極值。
植被的空間分佈也是野生動物的關鍵因素。河流的線性排列導致最大化的邊緣效應,增加物種豐富度,因為物種可以同時進入多種覆蓋 ( 或棲息地 ) 類型並利用兩者的資源 (Leopold1933) 。邊緣沿著多種棲息地類型發生,包括水體、河岸帶和內陸高地棲息地。
棲 息地之間老林連接建立森林高地那之間的連續性可環繞通過非森林領域。它們充當分散的飼養細胞係並促進植物和動物的再生。因此,連通性對於在景觀基礎上保留 生物多樣性和遺傳完整性非常重要。然而,棲息地的線性分佈或邊緣效應並不是所有物種棲息地質量的有效指標。利用棲息地島嶼而不是海洋島嶼進行的島嶼生物地 理研究表明,較大的棲息地島嶼既支持更多的鳥類,也支持更多的鳥類。更多的物種 (Wilson 和 Carothers1979) 。雖然連續廊道是最理想的,但下一個優選的情況是最小的碎片,即河岸植被的大塊地塊 ( � 島嶼 � ) ,大塊土地之間的空間最小。
爬行動物和兩棲動物
幾乎所有的兩棲動物 ( 蜥蜴,蟾蜍和青蛙 ) 都依靠水生棲息地進行繁殖和越冬。雖然受水的限制較少,但許多爬行動物主要在河道和河岸棲息地中發現。在亞利桑那州中西部發現的 63 種爬行動物和兩棲動物中有 36 種被發現使用河岸帶。在大盆地, 22 種爬行動物中有 11 種需要或喜歡河岸帶 (Ohmart 和 Anderson1986) 。
鳥類
鳥類是在河岸廊道中最常見的陸生野生動物。在全國範圍內,據報導,在一年中的某些時間裡,有 250 多個物種使用了河岸地區。北美築巢鳥類密度最高的是西南棉花棲息地 (CarothersandJohnson1971) 。在西南部的 166 種繁殖鳥類中,有 73 %喜歡河岸棲息地 (Johnson 等, 1977) 。
中西部河道的鳥類物種豐富,反映了廊道的植物多樣性和寬度。這些繁殖鳥類中有一半以上是在樹葉上捕食昆蟲的物種 ( 病毒,鶯 ) 或在地上尋找種子的物種 ( 鴿子,金鶯,雀喙,麻雀 ) 。接下來的是在地上或樹上覓食的食蟲物種 ( 鵝口瘡,啄木鳥 ) 。
Smith(1977) 報導,東南部森林棲息地的鳥類分佈與土壤濕度密切相關。 Woodcock(Scolopaxminor) 和鷸 (Gallinagogallinago) ,紅肩鷹 (Buteolineatus) ,連帽和 prothonotary 鳴鳥 (Wilsoniacitrina , Protonotariacitrea) 以及東南部的許多其他雀形目更喜歡河邊森林中的潮濕地麵條件灌叢餵食。沿著佛羅里達州的水道的柏樹和紅樹林沼澤擁有許多在東南部幾乎沒有的物種。
哺乳動物
河岸地區的覆蓋,水和食物資源的組合使它們成為大型哺乳動物的理想棲息地,如騾鹿,白尾鹿,駝鹿和麋鹿可以使用多種棲息地類型。其他哺乳動物在其部分或全部範圍內依賴於河岸地區。這些包括水獺 ( 水獺黃花 ) ,貓熊,浣熊、水狸、麝鼠、沼澤兔、短尾貂和水貂。
河岸地區為棲息地,水和大量蝙蝠物種的豐富獵物提供高密度覆蓋物,包括小棕蝙蝠 (Myotislucifugus) ,大棕蝠 (Eptesicusfuscus) 和蒼白蝙蝠 (Antrozouspallidus) 。布林森等人。 (1981) 美國大陸沿岸地區哺乳動物的幾項研究的結果列表他們得出結論,哺乳動物的數量一般在 5 到 30 之間,動物社區包括幾個鼠類,一個或多個大型哺乳動物,以及少數中小型哺乳動物。
胡佛和威爾斯 (Hoover and Wills , 1984) 報告了科羅拉多州三角葉林河岸林地中的 59 種哺乳動物,僅次於該地區其他八種森林覆蓋類型的松樹。在亞利桑那州中西部的土地管理局清單中發現的 68 種哺乳動物中有 52 種使用河岸棲息地。 Stamp 和 Ohmart(1979) 和 Cross(1985) 發現,與鄰近的高地地區相比,河岸地區的小型哺乳動物的多樣性和生物量更大。
在乾旱和半乾旱地區,河岸帶和周圍高地的哺乳動物物種多樣性和生產力之間的差異尤其明顯。然而,美國東部的低地大喬木對許多哺乳動物也有極高的棲息地價值。例如,低地大喬木支持的白尾鹿種群大約是高地森林相當面積的兩倍 (GlasgowandNoble1971) 。河道本身受某些動物活動的影響 (Forman1995) 。例如,水狸建造的水壩會導致水窪在河道或洪氾區內形成。水潭殺死了現有的大部分植被,儘管它確實為魚類和海水創造了濕地和開闊水域 gratorywaterfowl 。如果洪氾區中的適當木本植物稀少,則水狸將其切割活動擴展到高地,並且可以顯著改變河岸和河道。隨著時間的推移,水窪被替換由泥灘,成為一片草地,最終讓位給木本更迭 - 簡稱 AE ,下同階段。然后水狸經常在一個新的地方建造一座大壩,這個循環重新開始,只有一個空間位移。
沿河道的水狸築壩序列可能對水文,沉積和礦物質養分產生重大影響 (Forman1995) 。 來自暴雨的水被阻止,從而提供一些防洪措施。淤泥和其他細小沉積物積聚在水窪中,而不是在下游清洗。通常形成濕地區域,地下水位於大壩上游。水窪結合了緩 慢河流動,接近恆定的水位和低濁度,支持魚類和其他水生生物。鳥類可能廣泛使用水狸水窪。與洪氾區的典型波動不同,濕地也有相對恆定的地下水位。砍伐樹木 的水狸減少了榆樹、水曲柳等種類的豐富度。但增強了快速發芽的物種的豐度,例如赤楊,柳樹和楊樹。

圖 2.31 :植物垂直複雜性。複雜性可能包括許多植被層。
2D-2 水域生態系統
河 流的生物多樣性和物種豐度取決於可用棲息地的多樣性。自然運行、穩定的河流系統有利於棲息地的多樣性和可用性。這是河流穩定的主要區域之一,自然功能的恢 復一直是河流廊道恢復活動中的重要內容。河流的橫截面形狀和尺寸、坡度和限制、河床沉積物的細微性分佈,甚至其平面形狀都會影響水生生物的棲息地。在干擾 較小的情況下,狹窄、陡峭的壁橫截面比較寬的橫截面 ( 側邊較陡 ) 為棲息地提供的物理區域更少,但與較寬、較淺的河 流廊道相比,在深深潭中可能提供更豐富的生物棲息地。陡峭、受限的河流是一種高能環境,可能會限制棲息地的發生、多樣性和穩定性。許多陡峭、快速河流動的 溪流都是高價值的冷水鮭魚溪。無側限系統頻繁氾濫,可促進河岸棲地的發展。棲地隨河流蜿蜒而新增。河床中均勻沉積的大小比具有許多粒徑的河床提供的潛在棲息地多樣性更小。

圖 2.32 :河流系統及其棲息地子系統的層次結構。近似線性空間尺度,適用於二級序或三級序溪流。
2D-2.1 水域棲息地
水域環境總尺度,即水系本身,是以數十公里為組織量測的,而支流集水區或上、中、下游河段是以數百公尺為單元量測的, Reach 河段是以數十公尺為單元量測的。一個河段系統包括河彎、水瀑、急流、湍瀨 / 深潭等序列、深潭 / 湍瀨序列或其他類型的河床形式的組合,每種形式的規模都可能小於或等於 10 公尺。
在小尺度上。這些小棲地的例子包括樹葉或樹枝狀碎屑、鵝卵石或其他粗石塊上的泥砂或淤泥、巨石上的苔蘚或細礫石斑塊。陡坡通常在河流中形成湍瀨 / 水潭序列,特別是在鵝卵石,巨石和基岩河流中。每個湍瀨木石堆都充當微型級穩定結構。
湍瀨和深潭共同協作,以分配這些陡峭傾斜系統中可用的多餘能量。它們還為可用的棲息地增添了多樣性。不太陡峭的斜坡上的鵝卵石和碎石河流形成了水潭 / 湍瀨序列,這也增加了棲息地的多樣性。深潭為魚類提供空間,覆蓋和營養,它們為魚類在暴風雨,乾旱和其他災難性事件期間尋求庇護提供了場所。許多鮭魚物種的上游遷移通常涉及通過淺層區域的快速移動,然後在較深的水潭中休息。
表 2.11 :河流濕地功能。 Source: Brinson et al. 1995.

2D-2.2 河流濕地
河 道河流復育計畫可能包括復育濕地,如河流灘地植被系統或河岸濕地。這裡提供了濕地的一般性討論。河道河流復育計畫應設計用於保護或復育相關濕地的功能。濕 地是一種生態系統,它依賴於基質表面處或附近的恆定或反复的淺淹沒或飽和。濕地的最低基本特徵是在地表或附近經常性,持續淹沒或飽和,以及反映經常持續淹 沒或飽和的物理,化學和生物特徵。濕地的常見診斷特徵是含水土壤和水生植物植被。這些特徵將存在,除非物理化學,生物或人為因素已經消除或阻止其發展 (National Academy of Sciences1995) 。濕地可能發生在河流,河岸區和河道的洪氾區。河岸地區或區域可能包含濕地和非濕地。濕地是陸地和水生系統之間的過渡,其中地下水位通常在地表或附近,或者土地被淺水覆蓋。
對於植被濕地,水創建有利於水生植物的生長條件 - 植物在水或在子生長施特拉特即至少週期性缺陷的氧作為水含量過多的結果 (Cowardin 等人 1979) ,並促進土壤被水滲透飽和,土壤上層形成厭氧菌生長條件。
濕地功能包括魚類和野生動物棲息地,蓄水,沉積物捕獲,洪水災害減少,水質改善 / 污染控制和地下水補給。長期以來,濕地一直被認為是受威脅和瀕臨滅絕的魚類和野生動物物種的高生產力棲息地。濕地為聯邦政府列為受威脅或瀕危物種的 60 %至 70 %的動物提供棲息地 (Lohoefner1997) 。
聯邦地理數據委員會已採用美國魚類和野生動物管理局的美國濕地和深水棲息地分類 (Cowardin 等人, 1979 年 ) 作為濕地分類的美國標準。該服務的美國濕地清單 (NW1) 使用該系統執行其國會授權的角色,即識別,分類,繪製和數字化濕地和深水棲息地的數據。這個系統,這與科學的參考定義的美國美國科學院一貫定義的濕地,包括海洋,河口,河流,湖泊和沼澤系統。 NWI 還制定了在西部 22 個州的河岸棲息地進行分類和繪圖的協議。
Cowardin 分類下的河流水系包括河道中包含的所有濕地和深水棲息地,除了以樹木,灌木,持久性挺水植物,暫時性苔蘚或地衣和濕地原生物。
河岸帶地圖
河岸帶是兩種或兩種以上棲息地類型相遇時出現的最大值的典型例子。河岸棲息地的價值很重要。河岸地區植物群落連續並受到常年或間斷的地表和地下的水文特徵激河流和靜水水域 ( 河流,湖泊,排水方式 ) 。河岸地區具有以下一個或兩個特徵: (1) 與鄰近地區截然不同的營養物種 ;(2) 與相鄰區域相似的物種,但表現出更有活力或更強勁的生長形式。河岸地區通常在濕地和高地之間過渡。
河岸系統被細分為子系統、類、子類和支配類型是通過垂直高地限定在陸地側,在河道組 ( 包括天然和製造堤壩 ) ,或通過沼澤濕地。在辮狀分歧河流,河流濕地是有界的,通過形成在其內發生編織的凹陷的外部界限水岸。
在 該系統下,河道的植被河灘地濕地被歸類為綠洲。該沼澤系統被開發組植被濕地傳統上由這樣的名字沼澤,、草原和所謂的坑洞,也包括小的,淺的,永久或間歇水 體通常被稱為水窪。沼澤濕地可以位於向岸的湖泊,河流河道,或河口,對河流河灘地,在分離的集水區,或在斜坡上。它們也可能在湖泊或河流中出現。該沼澤系 統包括所有非潮汐由喬木,灌木,持續主導濕地突現,湧現的苔蘚和地衣,以及在潮汐地區發生的所有這些濕地,由於海洋來源鹽的鹽度低於 0.5ppt 。在沼澤系統是有界通過高地或通過任何其他四個系統。它們可能與非濕地河岸棲息地融合,水文條件不再支持濕地植被,或者在水文條件根本不支持濕地的情 況 下可能完全不存在 (Cowardin 等, 1979) 。
水文地貌 (HGM) 的方法是,用於進行濕地的功能評估分類成濕類似基團的系統。濕地根據地貌,水源和河流體動力學進行分類。這使得焦點可以放在一組濕地上,這些濕地的功能比沒有分類的情 況 更為相似。參考濕地用於開發參考評估濕地的標準 (Brinson1995) 。
在 HGM 方 法下,河流濕地發生在與河道相關的洪氾平原和河岸廊道中。主要水源是河道河流量或河道和濕地之間的地下連接。河流濕地通過地表和地下水流返回河道,地下水 補給和蒸發蒸騰而失去水分。在距離源頭最近的延伸段,河流濕地經常被坡道或窪地濕地所取代,其中河道床和河岸消失,或者它們可能與排水不良的平地和高地進 行交錯。通常是森林覆蓋,它們向下游延伸到陸地與河口邊緣濕地。橫向範圍是從河道的邊緣垂直於河灘地的邊緣。在一些景觀情 況 下,河流濕地可能在水文上更像坡地濕地,而在洪水平原很少或沒有洪水區的水源水中,坡地濕地可能位於河道附近。表 2.11 總結了 HGM 方法下河流濕地的功能。

圖 2.33 :河流生物群。溪流中發現的食物鏈關係。
表 2.12 :河流生物群通常觀察到的密度。

水生動物與植物
河流生物群通常分為七類 - 細菌,藻類,大型植物 ( 高等植物 ) ,原生生物 ( 變形蟲,鞭毛蟲,纖毛蟲 ) ,微小型無脊椎動物 ( 長度小於 0.5mm 的無脊椎動物,如輪蟲,橈足類,介重和線蟲 ) ,大型無脊椎動物 ( 長度大於 0.5mm 的無脊椎動物,如蜉蝣 , 石蠅,石蠶蛾,橈蝦,蚯蚓,蛤蜊和蝸牛 ) 和脊椎動物 ( 魚類,兩棲動物,爬行動物和哺乳動物 )( 圖 2.33) 。第 1 章中關於河流連續體概念的討論概述了河流中發現的主要生物群以及這些群落如何從高級序變為低級序。
未受干擾的溪流可能包含數量驚人的物種。例如,對德國一條小型河流佈雷滕巴赫 (Breitenbach) 的河流生物群進行了全面清查,在 2 公里河段 的範圍內發現了 1300 多種物種。可能在潛在恢復地點發現的藻類、大型無脊椎動物和魚類的清單可從州或地區的清單中獲得。這些河流生物群的密度如表 2.12 所示。
水 生植物通常由附著在永久性基質上的藻類和苔蘚組成。在基質適合的情況下可能發生有根的水生植被,並且高水流不會沖刷河流底部。維多利亞的維管植物床可能會 在某些地區生長,例如佛羅里達州的泉水流,其中存在水的透明度,基質,養分和緩慢的水流速度。通過河流不能輕易移動的基岩或石頭通常被苔蘚和藻類以及各種 形式的石頭覆蓋
浮游植物形態通常是有限的,但可能存在於河流流域包含湖泊,水窪池塘,河灘地水域或緩慢河流動區域的地方 (Odum)1971 年 ) 。 河流的底棲無脊椎動物群落可能含有多種生物群,包括細菌,原生生物、輪蟲、苔蘚蟲、蠕蟲、甲殼類動物,水生昆蟲幼蟲、貽貝、蛤蜊、磷蝦、螺類和其他形式的 無脊椎動物。水生無脊椎動物存在於河流中的多個微棲地中或其中,包括植物、植物枝幹碎片、岩石、硬質基質的間隙空間和軟基質 ( 礫石、沙子和淤泥 ) 。無脊椎動物棲息地存在於所有垂直地層,包括水面、水體、河床、河底地下和潛伏區內的深處。單細胞生物和微小無脊椎動物是河流中最多的生物群。但是,更大的無脊椎動物對群落 * 結構很重要,因為它們對河流的無脊椎動物總生物量有顯著貢獻。例如蜉蝣、蝸牛等吃附著藻類的初級消費者, 對河流中藻類和附生植物的豐度和分類組成產生顯著影響。同樣,大型無脊椎動物捕食者,如石蠅,可以影響無脊椎動物群落中其他物種的豐度 (Peckarsky 1985) 。
總的來說,微生物 ( 真菌和細菌 ) 和底棲無脊椎動物有助於分解從外部來源進入河流的有機物質,如落葉。一些無脊椎動物 ( 昆蟲幼蟲和微小甲殼動物 ) 充當碎食者,它們的進食活動將較大的有機落葉分解成更小的顆粒。其他脊椎動物從水中過濾較小的有機物質 ( 黑蠅幼蟲、一些蜉蝣若蟲和一些石蠅幼蟲 ) ,從表面刮下物質 ( 蝸牛、螺和一些石蠅和蜉蝣若蟲 ) ,或以沉積在基質上的物質為食 ( 雙翅目幼蟲和一些蜉蝣若蟲 )(Moss 1988) 。這些攝食活動導致有機物分解,而無脊椎動物組織則是其他消費群體 ( 如魚類 ) 賴以生存的。
底棲大型無脊椎動物 ,特別是水生昆蟲幼蟲和甲殼類動物,被廣泛用作河流健康和狀況的指標。許多魚類依靠底棲生物作為食物來源,或者直接在底棲生物上覓食,或者通過捕捉被移走並漂向下游的底棲有機物 (Walburg 1971) 。
魚類在河流生態系統中具有生態重要性,因為它們通常是最大的脊椎動物,通常是水生系統中的頂級捕食者。給定河流中魚類的數量和種類組成取決於地理位置,進化歷史以及物理棲息地 ( 水流、深、基質、湍瀨 / 水潭比,木障礙和底切岸 ) ,水質等內在因素 ( 溫度、溶氧、懸浮固體、營養物質和有毒化學物質 ) 和生物相互作用 ( 開發、捕食和競爭 ) 。
台灣地區大約有 160 種本地淡水魚類,其中有 63 種是只能在純淡水的河川、湖泊、埤塘中生活的初級淡水魚,另外的 100 餘種則是在河海交匯的河口及受海水漲潮所及的河段中所發現的魚類,包括 23 種可生活在淡水及河口一帶, 22 種可以雙向洄游,也就是一生中會經歷淡水、河口及海洋三種環境的魚種,以及 51 種只在河口及海洋而不會侵入純淡水域的種類。在這些魚類中,至少有 34 種是台灣的特有種,約佔全部魚種的 1/5 。中國大陸共有 700 多種的純淡水魚,另外能游入海內或河口的約有 70 種。其中種數量最多的鯉科有 400 多種,其中鯉形目共計佔總種數的 75% 左右。其次為 鮎 形目,兩者共佔全部魚種的 85% 以上。全球魚類共約 26,000 多種,其中有 41.2% 為淡水魚類,包括 33.1% 初級淡水魚, 8.1% 次級淡水魚, 0.6% 的兩向洄游魚種,它們所佔的比例如圖所示。換言之,全球淡水魚有將近 10,700 種,雖然總種數比海水魚少,但因全球淡水水域的體積只佔地球水體的 0.01% 不到,而卻有近半的全球魚種生活在淡水水域中,故淡水魚之相對豐度遠比海水魚要高的多。換另一種角度說,平均每種海水魚可棲息在 113,000 立方公里的水體,而淡水魚平均每種只有 15 立方公里的體積,高於海水約 7,500 倍。這主要的原因當然是因為陸域河川易受地理隔離阻絕,有助於特化的緣故。 ( 中央研究院台灣魚類資料庫 )
由 於控制溫度、溶氧、坡度、流速和底質的許多水文和地貌因素的變化,河流中的魚類物種組合將從源頭到出水口發生很大變化。這些因素結合在一起,決定了給定河 段內棲息地多樣性的程度。隨著坡度減小和河流尺寸增大,魚類物種豐富度趨於增加。由於坡度的增加和河流尺寸的減小,物種豐富度通常在小源頭河流處最低 (Hynes 1970 , Matthews and Styron 1980) 。此外,高梯度和與支流的聯繫減少,減少了新物種定居和進入的可能性。
由 於環境穩定性增加,潛在棲息地數量增加,定殖源數量增加或主要排水之間的聯繫增加,物種豐富度從中間到較低的河流河水流量遞增。當河流向下游行進時,深潭 和河道的數量會增加,可以增加細顆粒河床物質並促進大型植物的生長。這些環境允許魚類的存在更能容忍低氧和升高的溫度。此外,體形的範圍隨著具有較少的紡 錘形體形狀的那些物種的外觀而增加,其在生態學上適應於以降低的流水速度為代表的區域。在高級序河流或較大型河流中,底部基材通常以較細的泥砂、淤泥為代 表 ; 因此,食草魚類,雜食魚類和浮游生物可能因水生植被和浮游生物的可用性而增加 (Bond1979) 。魚類已經進化出獨特的覓食和繁殖策略,以便在不同的棲息地條件下生存。霍維茨 (1978) 檢查了 15 個不同水系魚類覓食同功群的結構,發現大多數魚類是底棲食蟲佔 33 %,而食魚魚類佔 16 %,食草魚類佔 7 %,雜食魚類佔 6 %,食浮游生物魚類佔 3 %,其他覓食同功群包含的物種較少。然而, Allan(1995) 指出,魚類經常改變棲息地,生命階段和季節的攝食習慣,以適應不斷變化的物理和生物條件。較小的上游河段中的魚往往是食蟲或刮食藻類,而隨著中下游河段條件的多樣性增加,覓食多樣的魚類數量和餵養策略的範圍會增加。
有一些魚類是遷徙的,長距離返回特定地點產卵。這些魚類可能表現出極大的耐力,向上游逆河流並向瀑布等障礙物遷徙。許多魚類必須在鹽水和淡水之間移動,需要很大的滲透調節能力 (McKeown1984) 。從海洋環境返回淡水流產卵的物種稱為溯河魚類,如鮭魚,從淡水河流往海洋產卵的物種稱為降海魚類,如鰻魚。還有些魚類配合季節、水溫、洪水周期,在主、支流間遷移,或在上、中、下游間遷徙。螃蟹也會配合季節到河口產卵。
物種通常可以稱為冷水或溫水,以及它們之間的等級,取決於它們的溫度要求 (Magnuson 等, 1979) 。 像鮭魚這樣的魚通常被限制在較高海拔或北方氣候,以較冷的高氧水為代表。這些物種往往是專家,具有相當狹窄的耐熱性和相當特殊的生殖要求。例如,鮭魚通常 通過在清潔礫石之上或之內沉積卵來產卵,所述清潔礫石由於間隙空間內的上升河流而保持氧化和無淤泥。生殖運動和行為受到微妙的控制熱變化與日長的增加或減 少相結合。因此,鮭魚種群極易受到許多形式的棲息地退化的影響,包括河流量,溫度和基質質量的改變。
許多淡水魚類數量正在下降,或種類滅絕。魚類科學家提出了應歸類為瀕危,受威脅或特別關注的魚類物種清單。該清單包含 364 種因其稀有性而需要保護的魚類。棲息地喪失是所列物種中約 90 %消耗的主要原因。該研究指出,在 25 %的州中發現了 77 %的魚類物種,其中西南部八個州的濃度最高。 Nehlsen 等。 (1991) 提供了 214 份來自加利福尼亞州,俄勒岡州,愛達荷州和華盛頓州的貧瘠太平洋鮭魚等。引為下降原因是魚道和遷移的改變由於水壩,相關聯的河流量減少與蓄水發電、農業、沉積和棲息地損失,由於記錄和農業,過度捕撈,並與其他魚負相互作用,包括非天然鮭魚和虹鱒魚。
當 地魚類數量的普遍減少導致目前對復育魚類棲息地質量和數量的廣泛興趣。復育活動經常集中在改善當地棲息地,例如圍欄或從河流中移除牲畜,建造魚類河道或安 置河道物理棲息地。然而,研究表明,在大多數情況下,成功是有限的或可疑的,因為重點太狹窄,並沒有解決物種生命週期所需的各種棲息地要求和資源的復育問 題。
河道修復工作者和其他人現在敏銳地意識到魚類在整個季節和生命週期中需要許多不同的棲息地來滿足餵養,休息,避免掠食者和繁殖的需要。例如,利文斯通和拉貝尼 (1991) 確定密蘇里州東南部傑克斯福克河中的少年小嘴鱸主要以沿岸植被中的小型大型無脊椎動物為食。植被不僅是食物來源,而且是掠食者的庇護區和更溫暖的棲息地,這些因素可共同優化生存和成長的機會 (Rabeni) 和 Jacobson1993) 。然而,成年小口鱸魚往往佔據更深的水潭栖息地,不同地點成蟲的數量和生物量都歸因於這些特定的深水棲息地 (McClendon 和 Rabeni1987) 。 Rabeni 和 Jacobson(1993) 建議了解這些特定的棲息地,並結合對河流水力學和地貌學的理解形成和維持他們的關鍵是發展成功的河流復育項目。
由於許多生態,經濟和遊憩因素,對魚類群落復育的重視正在增加。 1996 年,大約有 3500 萬 16 歲以上的美國人參加休閒垂釣,導致支出超過 360 億美元 (Brouha1997) 。大部分活動都在河流中,這證明瞭河道河流復育計畫的合理性。
雖 然魚類種群經常受到公眾的極大關注,但保護其他水生生物群也可能是河流量復育的目標。淡水貽貝,其中許多物種受到威脅和瀕臨滅絕,往往是特別令人關注的問 題。貽貝對棲息地干擾非常敏感,顯然受益於完整,管理良好的河道。美國中南部擁有世界上最多樣化的貽貝。貽貝生態學也與魚類生態密切相關,因為魚類作為貽 貝幼蟲 (glochidia) 的寄主 ) 。他們面臨的主要威脅是水壩,導致直接棲息地喪失和剩餘棲息地破碎,持續沉積,殺蟲劑和引入外來物種,如魚類和其他貽貝物種。
水生系統中的非生物和生物相互關係
河流生物群的大部分空間和時間變化反映了非生物和生物因素的變化,包括水質,溫度,河流量和河流速,基質,食物和營養物的可用性以及捕食者 - 獵物關係。這些因素影響水生生物的生長,存活和繁殖。
水流條件
從 上游到下游的水流將河流與其他生態系統區分開來。水流的空間和時間特性,如快與慢、與深淺、湍急與平順、洪水與低緩,被描述本章前面。這些河水流動特徵可 以影響許多河流物種的微觀和宏觀分佈模式。許多生物都與河水流速敏感關連,因為它代表了運送食物和營養素的重要機制,但也可能會限制生物體保留在河段中的 能力。一些生物也響應河流動的時間變化,這可以改變河流水道的物理結構,或者增加死亡率,改變可用資源和破壞物種之間的相互作用 (Resh 1988 , Bayley 1992) 。
河流中的流速決定了浮游生物形態是否能夠自我發展和維持。河流中的水流越慢,岸邊和底部的生物群的組成和構造越接近於靜水狀態,浮游藻類及浮游微生物繁殖更加有利。高河水流量 ( 洪水氾濫 ) 是定時遷移和某些魚類產卵的線索。高河水流量還可清潔和搬運河床物質和沖刷池。極低的河水流量可能會限制幼魚的生產量,因為這種流量經常發生在幼魚的繁殖和生長期 (Kohler 和 Hubert1993) 。
水溫
河流水作為環境空氣溫度調節的功能,水溫可在河流系統內與河流周邊環境顯著變化,緯度、水的來源和太陽輻射 (Ward1985 , Sweeney1993) 。溫度控制著冷血水生生物的許多生化和生理過程,因為它們的體溫與周圍的水相同 ; 因此,水溫決定生長,發育和行為模式方面具有重要作用。例如,水生昆蟲通常在較溫暖的河流部分或在溫暖的季節中生長和發育得更快。如果地點之間的熱差異顯著 ( 例如,沿緯度或海拔梯度 ) ,某些物種有可能在較溫暖的地點每年完成兩代以上的繁殖 ; 這些相同的物種每年在較冷的地點完成一代以下的繁殖 (Sweeney1984 , Ward1992) 。藻類和魚類的生長速率似乎以類似的方式響應溫度變化 (Hynes1970 , Reynolds1992) 。溫度與生長,發育和行為之間的關係可以強大到足以影響某些物種的地理範圍 ( 表 2.13) 。
水溫對於溶氧濃度有直接的影響,水溫是決定淡水流中魚類分佈的最重要因素之一,並受當地條件 ( 如陰影、深度和水流 ) 的影響。許多魚類只能忍受有限的溫度範圍。像鮭魚、鱒魚與 鯝 魚這樣的魚在冷水流中占主導地位,而大口黑鱸、小嘴鱸、鯉魚、翻車魚和鯰魚等物種可能會出現在溫暖的河流中 (Walburg , 1971) 。
樹覆蓋效果
對於河流生態復育的目的,土地使用方式的移除河畔樹林遮蔽或減少河水流量,可能增加河道內溫度超過了魚類臨界的最大值 (Feminella 1984) 。因此,維持或恢復正常溫度狀態可能是河流管理者的重點。河岸植被是影響河流光照和溫度衰減的重要因素 (Cole1994) 。直射陽光可以顯著加熱河流,特別是在低流量的季節。在這種情況下,山地河流在進入森林砍伐區時會迅速變暖,但當河流重新進入森林茂密區時也可能會有所降溫。有時候河水溫度降低可歸因於較冷的地下水的河流入。缺乏覆蓋也會影響冬季的河流溫度。 Sweeney(1992) 發現 2 至 6 � C 的 溫度變化通常會改變研究物種的關鍵生活史特徵。河岸植被緩衝帶已被證明,對於防止水溫,營造魚類棲息地方面很有效。溫度控制所需的確切緩衝區寬度因地點而 異,具體取決於河流方向,植被和寬度等因素。沿著較小的狹窄的水源,重新建立灌木,例如柳樹和赤楊,可以提供足夠的遮蔭和有機物碎屑,以復育河岸和水生生 態系統。種植和 / 或重建大樹,例如三葉楊,柳樹,梧桐和核桃 (Lowe1964) ,沿著較大的高級序下游河段河岸,可顯著改善了最魚類繁殖條件。太陽輻射是影響夏季水溫峰值的主要因素,遮蔭對小河流流域河流的整體溫度狀況至關重要。
溶氧
氧氣直接從大氣中吸收並通過植物光合作用進入水中 (Mackenthun , 1969) 。由於深度淺,大面積暴露於空氣和恆定運動,即使在光合作用下沒有氧氣產生時,河流體通常也含有豐富的溶氧供應。
適當濃度的溶氧不僅對保持水生生物的存活至關重要,而且對維持其繁殖,活力和發育至關重要。生物體在降低氧氣水平時會在維持物種 ( 競爭 ) 產生壓力 (Mackenthun1969) 。由於多種原因,溶氧濃度低於 3.0mg/L 時,已被證明會干擾魚類種群的生存 (Mackenthun1969 ,引用其他幾個來源 )( 表 2.14) 。
溶氧的消耗可導致包括魚類在內的多種水生動植物死亡。當生物和化學過程對氧氣的需求超過再氧化和光合作用輸入的氧氣時,魚就會死亡,從而導致魚類窒息。氧 耗竭通常與緩慢的水流、高溫、有根的水生植物的大量生長、藻類大量繁殖或高濃度的有機物質有關。
河流社區易受污染,也將減少溶氧供應 (Odum1971) 。決定水中氧含量的主要因素是溫度、壓力、水生植物的豐度以及水體與大氣接觸面的自然通氣量。水中 5mg/L 的溶氧水平將能維持大多數魚類的正常活動 (Walburg , 1971) 。冷水鱒魚河流的氧濃度需求範圍為 4.5 至 9.5mg/L(Needham1969) 。
酸鹼值
pH 值觀系到多種類群的水生生物生存,具有接近中性的氫離子活性 (pH7) 可提供水生系統中茁壯成長。 pH 值過低或酸性環境會抑制許多物種繁殖,降低物種多樣性和豐度 ( 圖 2.34) 。 pH 值變化的一個更廣泛認可的影響歸因於某些地區降雨的酸度增加,尤其是工業區和城市排放的下風側水體 (Schreiber1995) 。雖然鹼性土壤具有有限的緩衝能力,可以中和酸輸入情況。但是過多的酸雨仍可能導致逕流趨於略微酸性。
河流底質
河流生物群響應於受河床物質的影響,也波及許多生物和非生物變量。例如,在單一河流範圍內的障礙物 ( 樹木堆積 ) 、泥砂、岩塊和卵石中發現的大型無脊椎動物群落,可以觀察到物種組成和豐度的差異 (Benke 等, 1984 , Smock 等, 1985 , Huryn 和 Wallace , 1987) 。這種對不同基質相關條件的偏好有助於在較大的空間尺度上觀察到的模式。在河濱、山麓和山澗中發現了不同的大型無脊椎動物組合 (Hackney 等, 1992) 。
河床基本物質構成了水與水生系統的表面界面。所謂 � 潛河流區 � 是河床下方飽含水的軟泥層,並且可從河流廊道的下方和橫向延伸的深度,一個非常大的地下水滲流的環境層範圍。許多河流的洪氾平原河段都有一個具有明顯地表水 / 地下水相互作用的 � 潛河流區 � ,甚至可能造成伏流景觀 ( 河流水脈進入地下,隔一段距離再湧出地面 ) 。 Naiman(1994) 等人討論了太平洋西北地區河流周圍的潛河流帶的範圍和連通性。他們推測,作為一個從低級序 ( 小 ) 河流移動到高級序 ( 大 ) 河 流,潛河流區的重要性和連續性先增大後減小。在小河流中,潛河流帶僅限於小型洪氾平原,草地和河流段,其中粗糙的沉積物沉積在基岩上。該潛河流區一般是不 連續的。在具有更廣泛氾濫平原的中游河段中,潛河流區的空間連通性增加。在高級序的大河流中,潛河流區的空間範圍通常最大,但由於與河流活動相關的特徵, 如牛軛湖和截流河道,以及由於局部,中間和區域地下水系統的複雜相互作用,它往往是高度不連續的 (Naiman 等, 1994)( 圖 2.35)) 。
河床基質由各種物質組成,包括粘土、泥砂、礫石、鵝卵石、大塊石、有機物和樹木碎片。基材形成固體結構,改變表面和間歇河的流動模式,影響有機物質的積累,並提供生產、分解和其他過程 (Minshall1984) 。泥砂和淤泥通常是支持水生生物,並支持最少物種和個體的最不利的基質。扁平或碎石塊基質具有最高的密度和最多的生物量 (Odum1971) 。如前所述,河床基質的尺寸、異質、相對於高流量和基本流量的穩定性並且耐久性在河流內變化,取決於顆粒尺寸,密度和河流動能。河床無機基質往往在上游比下游粒徑更大,在湍瀨淺灘段的石塊比深水潭中的砂礫更大 (Leopold 等人, 1964) 。同樣,樹木枝幹碎片的分佈和功能,隨著河流的規模大小而變化 (Maser 和 Sedell1994) 。
在森林覆蓋的河流流域,以及河岸廊道中有大量樹木的河流中,落入河流的大型木質碎片 ( 枝、幹、葉、果 ) 可以增加河床基質和水生棲息地或範圍的多樣性。木質碎屑壩形成湍瀨淺灘,將泥砂、有機物沉積物截留,並經常在下游側形成沖刷坑。侵蝕的河岸通常發生在碎片堵塞的邊界。
有機物質傳輸
河流中的代謝活動取決於本地、異地和上游的食物和營養來源 (Minshall 等, 1985) 。 本地原生的有機物,如藻類和水中微生物,從該河段本地生產,而異地物質,如樹幹、枝葉、動物殘骸和溶解的有機碳,來自其他河段或其他支流或河岸兩側陸域。 上游物質可以是本地原生的或異地輸入的,並且通過河流動輸送到下游位置。季節性洪水為河道提供了有機物質的異地輸入,並且還可以顯著提高有機物質的分解速 率。
河流的初級生產力的作用,可能根據地理位置、河流大小和季節大不相同 (Minshall1978 年 ) 。 河流連續性概念推測,初級生產力的最小重要性的意義,因為河流大小的增加和河岸植被沒有陰影河源河流,但增加更長時間限制光線進入河流體附生生物。許多研 究人員已經證明,河流源流河段溪流窄小,且大多位於森林茂密覆蓋的地區,陽光不易透射到河流,水體生長藻類很少,初級生產力很弱。二至三級序河流,河谷較 寬,河畔樹木不能完全遮蔽水面,河流大塊石或砂洲之間也積留泥砂,長滿草類、矮灌木,初級生產力很高。河流的植物群可以是高山河流中的矽藻,也可以是低緩 河流中的茂密大型植物。
低流變帶 Hyporheic zone
低流變帶是在河床下面和旁邊的沉積物和多孔空間的區域,在這裡淺層地下水和地表水混合在一起。該區域的水流動力學和行為被認為對地表水 / 地下水的相互作用以 及魚類的產卵以及其他過程都很重要。作為一種創新的城市水管理實踐,低流變帶可以由工程師設計並進行積極管理,以改善水質和河岸生境。

圖 2.34 :酸雨對某些水生物種的影響。當酸度新增 (pH 值降低 ) 時,湖泊和溪流,一些物種消失了。

圖 2.35 : Hyporheic 潛河流區。 河流底棲生物群所經歷的不同棲息環境。
表 2.13 :所選魚類生長的最高周平均溫度和短期最高溫度。 Brungs and Jones 1977.

表 2.14 :與鮭魚和非鮭魚類水域中對魚類影響相關的溶解氧濃度 (mg/L) 匯總。 USEPA 1987.

正如前面第 2.C 節所討論的,向河流中注入氮和磷可以提高藻類和水生植物的生長速度,這一過程被稱為優養化 ( 富營養化 ) 。魚類和其他物質的過度分解會導致水體中氧氣和其他物質的消耗。
湖泊和水庫的優養化是以浮游植物生物量 ( 通常以浮游植物葉綠素濃度為代表 ) 間接衡量的。然而,在較小的河流中,浮游植物的生物量通常不是植物生物量的主要部分,這是由於能量流週期和高基質體積比有利於河底附生植物和大型植物的發育。河流富營養化可導致在流量 減 少和溫度升高時過度的藻墊和氧氣消耗 ( 圖 2.36) 。
此外,在環境氮、磷濃度明顯較低的溪流中,植物會過度生長,因為溪流促進植物細胞表面營養物質和代謝廢物的有效交換。
在許多溪流中,陰影或濁度限制了藻類生長所需的光照,生物群高度依賴外來有機物,如周圍流域產生的樹葉和樹枝。一旦樹葉或其他外來物質進入河流,它們就會發生迅速的變化 (Cummins1974) 。可溶有機化合物,通過浸出去除。細菌和真菌隨後在葉子上定殖並代謝它們作為碳纖維微生物生物量的存在新增了葉片的蛋白質含量,這最終代表了粉碎無脊椎動物的優質食物資源。
微生物分解和無脊椎動物粉碎 / 刮 削的結合降低了有機物的平均粒徑,導致碳的損失,既作為呼吸的二氧化碳,也作為較小的有機顆粒被輸送到下游。這些較細的粒子,從一個流段流失,成為下游部 分的能量輸入。營養物質和有機質在洗劑系統中的單向運動由於暫時保留、儲存和利用葉包、堆積的碎屑、無脊椎動物和藻類中而 減 慢。有機質加工與初級生產力有著相似的營養依賴關係。
葉 片和其他形式的有機物的分解可以受到氮或磷的限制,預測的氮磷比與藻類和附生植物的生長相似。葉片分解是由微生物分解、無脊椎動物切碎和物理分餾的順序組 合發生的。葉片和有機質本身的蛋白質含量通常較低。然而,由於微生物生物量中蛋白質和脂類的積累,細菌和真菌對有機物的定殖新增了氮和磷的淨含量。這些化 合物是水生無脊椎動物的主要營養來源。腐爛的有機物是溪流中營養物質的主要儲存成分,也是食物網中能量和營養物質轉移的主要途徑。最終,保留和利用的效率 以魚類生物量的形式反映在食物網的頂部。
有機體通常對本地、外來和上游來源的可用性變化作出反應。例如,與以封閉的樹冠和積累的葉子作為主要食物資源的溪流相比,食草動物在具有開放的河岸樹冠和高藻類生產力的溪流中相對更常見 (Minshall 等人。 1983 年 ) 。在同一條河流的縱向上也可以觀察到類似的模式 (Behmer 和 Hawkins 1986) 。
陸域與水域生態系統組成
前 面的部分介紹了塑造河道的生物組成部分和功能過程。為了簡單和易於理解,分別討論了陸地和水生環境。不幸的是,這通常與環境河流復育計畫採取相同的方法, 分別在高地,河岸區或河道河道上進行。必須將河道視為單個功能單元或生態系統,其中組件之間存在大量連接和交互。成功的河道復育不能忽視這些基本關係。
植被的結構和功能在所有尺度上都是相互關聯的。它們也與生態系統動態直接相關。特定的植被類型可能具有特徵性的再生策略 ( 例如,火災、樹木間隙 ) ,這些策略始終在景觀中保持這些類型。同樣,由於大量植物碎片堆積堵塞或水狸築壩,某些地形設置可能比其他設施更容易受到水文和相關植被結構的周期性劇烈變化的影響。然而,在河道生態系統的背景下,一些最基本的動態相互作用與河流洪水和河道遷移有關。
許多生態系統功能受植被結構特徵的影響。在未開發的河流流域,水和其他物質的運動受到植被和碎屑的調節,營養物質以複雜的模式被動員和保存,通常導致陸地和土壤之間的平衡相互作用。
水生系統
如果去除生物量,農業,牲畜草食者,開發和其他土地利用來改變植被的特徵和分佈,改變水、沉積物和養分的河流動模式,系統組成部分之間的相互作用將變得不 那麼有效。當引入過量營養素和合成毒素,土壤干擾和類似影響加劇這些問題時,這些問題會變得更加明顯。
在大多數未受干擾的洪氾區內,河流遷移和洪水是植物群落內部和之間結構和組成變化的主要來源 (Brinson 等, 1981) 。 雖然河流對植物群落產生了複雜的影響,但植被直接影響河流水系的完整性和特徵。例如,根系結合堤岸沉積物和中度侵蝕過程,洪氾平原植被減緩了岸上河流動, 引起沉積物沉積。落入河道的樹木和較小的木質碎片會阻礙流動,誘導某些點的侵蝕和其他地方的加強沉積、改變水潭分佈、有機物質的運輸,以及許多其他過程。 洪氾區與河流高度互動的的穩定,可以直接影響河道生態系統結構和功能的基本過程,從而間接影響周圍景觀的特徵。
陸域系統
在大多數情況下,最明顯的植被功能是影響魚類和野生動物的功能。在景觀層面,本土植被覆蓋的碎片化已經顯示出對野生動物的顯著影響,棲地條件將偏向於機會主義物種 ( 掠食動物 ) ,而不是那些需要大量鄰近棲息地的物種 ( 青蛙、溪蟹、石蛾 ) 。 在某些系統中,廊道連續性的相對較小的中斷,可能對動物運動或河流條件支持某些水生物種的適宜性產生重大影響。與本地生態系統結構不搭配的組合,或不適當 的廊道可能同樣具有破壞性。狹窄的廊道可能使邊緣棲息地吸引普通常見的物種,如築巢土蜂和掠食者;排除需要高隱密性的弱勢物種。或者廊道已經建立了柵欄、 圍牆、陡坡、硬鋪面、堤防等物種跨越的障礙,它們將破壞區域動物組合的完整性。
一 些與仰賴河岸生存的物種必須與河岸邊緣棲息地緊密相連,這些地帶通常有相當連續的茂密樹冠。如果這些植被種類太過單一、穩定,沒有新的樹木進入既有植物 群,這些茂密相連植被的老樹最終會萎縮,從而形成更小的棲息地。適當的干擾,將能活化老樹植被的生態條件。物種來源能夠獲得足夠的補充和斑塊大小重組,將 使許多物種受益。出於類似的原因,許多與河岸有關的猛禽如常見的黑鷹,灰鷹,禿鷹,貓頭鷹,都有助於河岸棲息地的活力。獲得足夠不同樹木補充,將使當地物 種組合長期受益。
與 該主題相關的一些方面已被討論為其他部分下的生態系統組成部分和功能。最早的關於河岸棲息地破碎對繁殖鳥類影響的研究結果發表在西南地區。其他研究人員的 後續研究發現了類似的結果。基本上,北美西南部的棉花林畫廊森林支持最高濃度的非殖民地為北美築巢的鳥類。這些河岸森林的破壞和破碎減少了物種的豐富度, 並導致築巢對 / 英畝的數量與成熟樹木 / 英畝的數量之間幾乎呈直線關係。後來的研究表明,河岸地區與遷徙鳥類的管道同樣重要 (Johnson 和 Simpson1971 , Stevensetal.1977) 。
在 考慮復育河岸棲息地時,必須考慮相鄰棲息地的狀況。生物學家發現河岸生態系統,特別是水濱邊緣帶,被親水鳥類、水生昆蟲、兩棲爬蟲類廣泛使用。並且發現, 一些濱水鳥類利用相鄰河畔的生態系統築巢、覓食。也現較小體形的物種繁殖,很多會利用河流廊道陸域地帶進行。較大體形的物種通常在鄰接河岸生態系統 ( 河廊 ) 外側陸域覓食。較大型的掠食物種 ( 如猛禽 ) 可能會從河岸生態系統中覓食數平方公里,但是仍然非常依賴河流廊道的陸域 (Lee 等, 1989) 。
由於峽谷效應產生的更多中間條件,峽谷及其伴隨的河岸植被作為沿著海拔梯度 ( 例如,夏季和冬季範圍之間 ) 的動物短程移動的廊道。沿著河岸帶發生的長程遷移物種,包含遷移鳥類、蝙蝠、魚類。河岸帶也是季節候鳥的中途棲息地 (Stevens 等, 1977) 。木本植被通常很重要,不僅對大多數河岸生態系統,而且對鄰近的水生甚至高地陸域生態系統都很重要。但是,在嘗試改變棲息地之前,必須建立明確的管理目標。將所有特定生態系統復育到 � 原始狀態 � 可能是不可能的。特別是上游集水區條件已經被大量修改,例如構築大壩或其他引水工程、山坡地開發等,更難恢復。即使大壩、灌溉等能完全恢復,它也可能無法實現複雜的生態系統復育目標。
2E 河流生態功能和動態平衡
■ 棲地: 環境的空間結構,允許物種生存、繁殖、覓食、移動和庇護。
■ 屏障: 阻擋物質、能量和生物的停止前進。
■ 導引: 傳輸物質、能量和生物有機體的能力。
■ 篩濾: 過濾物質、能量和生物有機體的選擇性滲透。
■ 來源: 物質、能量和生物體輸出超過輸入的機制,如物種繁殖場、湧泉。
■ 匯聚: 水、能量、生物和物質的輸入超過輸出的關鍵區,如水槽匯聚。

圖 2.37 :關鍵生態系統功能。六個功能可以概括為一組在各種設定中重複出現的基本的、共同的主題。
在 過去的兩章中,本書涵蓋了河道結構以及河道中發生的物理,化學和生物過程。該信息顯示了河道如何作為生態系統發揮作用,因此,必須如何理解這些特徵結構特 徵和過程,以便有效地復育河道功能。事實上,重建結構或復育特定的物理或生物過程並不是復育力求實現的唯一目標。復育旨在重建有價值的功能。專注於生態功 能使復育工作成為重建自我維持系統的最佳機會。可持續性的這一特性是功能性聲音的分離河流,自由地為人類和自然環境帶來許多好處,來自受損的水道,無法維 持其有價值的功能,並可能仍然是昂貴的長期維護負擔。
第一章第一節 A 節強調矩陣、斑塊、廊道和馬賽克是區域尺度上最基本的物理結構構建塊。生態功能也可以概括為一套基本的、共同的主題,在無限的環境中重現。這六個關鍵功能是棲息地、導管、篩檢程式、屏障、源和水槽 ( 圖 2.37) 。在這一部分中,我們將從這些關鍵的生態功能出發,重新審視過去兩章的過程和結構描述。對於流廊函數的操作,兩個屬性特別重要:
連通性這是衡量廊道或矩陣空間連續性的一種度量 (Forman 和 Godron 1986) 。該屬性受廊道中的間隙或斷裂以及廊道與相鄰土地使用之間的影響 ( 圖 2.38) 。一條河流廊道在自然群落之間具有高度連通性,促進了寶貴的功能,包括材料和能源的運輸以及動植物的移動。
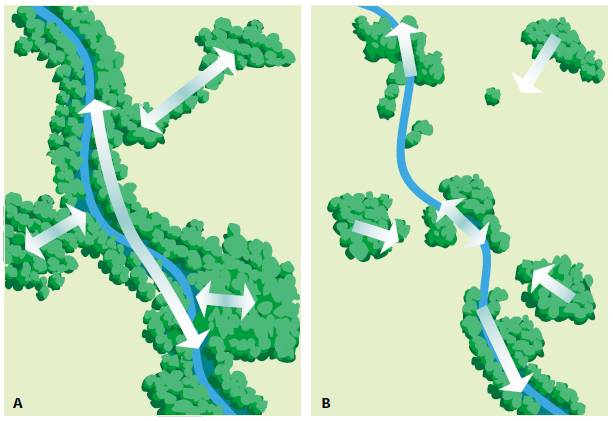
圖 2.38 :具有 (A) 高連通度和 (B) 低連通度的景觀。一個連通的景觀結構通常比一個支離破碎的景觀具有更高的功能水準。
河流廊道寬度,指河流及其鄰近植被覆蓋區之間的距離。影響寬度的因素有邊緣、群落組成、環境梯度和鄰近生態系統的干擾效應,包括人類活動的影響。寬度的示例性度量包括平均尺寸和方差、狹窄數量和不同生境要求 (Dramstad 等人 1996 年 ) 。
寬 度和連通性在整個河流廊道的長度之間相互作用。道路寬度隨河流長度而變化,可能有間隙。廊道之間的間隙中斷並減少連接。評估連線性和寬度可以為設計減輕干 擾的恢復措施提供一些最有價值的見解。下面的小節討論了每個函數以及與連接和寬度的一般關係。最後一部分討論了動態平衡及其與河道恢復的相關性。
棲息地是指植物或動物 ( 包括人 ) 通常生活、生長、覓食、繁殖或以其他方式存在於其生命週期的任何部分的區域。生境為生物體或生物體群落提供了必要的生命要素,如空間、食物、水和住所。
以下小節討論了每個功能以及與連接和寬度的一般關係。最後一小節討論了動態平衡及其對河流廊道復育的提升。
邊緣和內部棲息地
兩個重要的棲息地特徵是邊緣和內部 ( 圖 2.39) 邊 緣是不同生態系統之間相互作用的關鍵線。內陸棲息地通常是更穩定,更有庇護的環境,生態系統可能在較長時間內保持相對相同。邊緣棲息地暴露於高度可變的環 境梯度。結果是與觀察到的內部棲息地不同的物種組成和豐度。邊緣對於擾亂內部棲息地的過濾器很重要。邊緣也可以是各種各樣的動植物群。
邊緣和內部是與尺度無關的概念。被稱為內陸森林物種的較大的哺乳動物可能需要距離森林邊緣數英里才能找到所需的棲息地,而昆蟲或兩棲動物可能對腐爛的原木 下的微棲地的邊緣和內部敏感。因此,河道的邊緣和內部取決於所考慮的物種。作為狹長的狹窄生態系統,包括陸地 / 水界面,並且通常包括自然 / 人造邊界以及高地邊緣,河道具有豐富的邊緣,這些對其生物群具有顯著影響。
不同的植物和動物物種都喜歡邊緣和內部,並且將邊緣或內部視為始終 � 壞 � 或 � 好 � 的棲息地特徵是不合適的。在某些情 況 下可能需要保持或增加邊緣,或在其他情 況 下有利於內部棲息地。然而,一般而言,人類活動傾向於增加邊緣並減少內部,因此更常見的是復育或保護內部,這需要特定的管理行動。
河道邊界的邊緣棲息地通常具有更高的太陽能,降雨,風能和相鄰生態系統的影響。河道邊緣的環境梯度差異導致多樣化的植物和動物群落與相鄰的生態系統相互作用。當內部棲息地的數量最小時,邊緣的影響更加明顯。 內部棲息地從元素的周邊進一步發生。內部是代表由比那些在生態系統的邊緣發現更穩定的環境輸入。棲地內陸區陽光,降雨和風力影響較小。許多敏感或稀有物種依賴於較少受干擾的環境來維持生存。因此,它們只能容忍 � 內部 � 棲息地條件。創建這些內部條件所需的周長距離取決於物種的要求。

圖 2.39 :林地邊緣和內部棲息地。室內植物和動物與那些喜歡或容忍邊緣變化的植物和動物有很大的不同。
棲息地功能
棲息地是指植物或動物 ( 包括人 ) 通常生活、生長、餵養、繁殖或以其他方式存在於其生命週期的任何部分的區域。棲地為生物體或生物體群落提供了必要的生命要素,如空間、食物、水和住所。
在 通常由河流廊道提供的適當條件下,許多物種可以利用廊道生活、尋找食物和水、繁殖和建立可行的種群。穩定生物群落的一些措施是種群規模、物種數量和遺傳變 異,這些變化隨時間的推移在預期的範圍內波動。不同程度上,河流廊道對這些措施產生了建設性的影響。廊道作為棲息地的價值增加了,因為廊道經常連接許多小 棲地斑塊,從而創造出更大、更複雜的棲息地,野生動物種群更大,生物多樣性也更高。
棲地功能在不同的尺度上有所不同,對不同棲地功能發生的尺度的瞭解將有助於恢復倡議的成功。例如,對大尺度棲地的評價可能會注意到生物群落的大小、組成、連通性和形狀。
在景觀尺度上,矩陣、斑塊、馬賽克和廊道的概念常常涉及到描述大面積棲息地的概念。河流廊道和主要河谷一起提供了大量的棲息地。北美的飛道包括河流和河流廊道棲息地的例子,這些棲息地是候鳥在景觀上開發的,在區域尺度上。
河流廊道和其他類型的自然植被廊道,也可以為遷徙的森林和河岸物種提供它們在遷徙中途停留期間的首選休息和餵養棲息地。眾所周知,黑熊等大型哺乳動物需要 大而連續的野生地形作為家系,在該國許多地區,寬闊的河流廊道對於將較小的斑塊連接到足夠大的領土至關重要。
從 不同的角度來考察流域內的棲地功能。流域內的棲息地類型和模式非常重要,與相鄰流域的連通性模式也非常重要。流域上游河流廊道的植被有時與分水嶺以外的相 鄰流域和廊道的植被斷開。當陸地或半水上河流廊道群落在水源處連接時,這些連接通常有助於提供流域以外的適當替代棲息地。
評 估河流廊道和較小尺度的棲息地功能也可以從斑塊和廊道的角度進行,但比景觀和流域更詳細。在地方尺度上,廊道內各種棲地之間的過渡也會變得更為重要。河流 廊道通常包括兩種一般類型的棲地結構:內部和邊緣棲地。棲息地多樣性是由一條包括邊緣和內部條件的廊道增加的,儘管對於大多數河流,廊道寬度不足,無法為 較大的脊椎動物提供大量的室內棲息地,例如森林內部鳥類物種。因此,增加內部棲地有時是流域規模恢復的目標。
廊道尺度上的棲地功能受連通性和寬度的影響較大。河流廊道沿線和跨河廊道的連通性和寬度的增加通常會增加其作為棲息地的價值。河谷形態和環境梯度 ( 如土壤濕度、太陽輻射和降水的逐漸變化 ) 可能導致植物和動物群落的變化。與狹窄、均勻或高度破碎的廊道相比,更多的物種在河流廊道內廣泛、連續和多樣的原生植物群落中找到合適的棲地條件。
但是,當嚴格應用於流通道時,這可能不是真的。例如,一些狹窄和深切的河流提供了對瀕危輸卵管炎至關重要的熱條件。
廊 道內的棲地條件因氣候和微氣候、海拔、地形、土壤、水文、植被和人類用途等因素而異。在規劃恢復措施方面,廊道寬度對野生動物尤為重要。例如,在規劃維護 特定野生動物物種時,廊道的尺寸和形狀必須足夠寬,以包括足夠合適的棲息地,使該物種能夠填充河流廊道。廊道太窄,可能會像廊道上的一個完整的縫隙一樣, 為某些物種的移動提供障礙。在局部尺度上,大量的木質碎片被卡在河道中,可以對河流和鄰近的河岸造成形態變化。
深潭可以從流經河流的原木下游形成,上游和下游的水流特性都會發生變化。河流中的大型木本碎片形成的結構改善了大多數魚類和無脊椎動物的水生棲息地。
河岸林除了邊緣和內部棲息地外,還可能在其樹冠層、小喬木、灌木層和草本層中提供垂直棲地多樣性。在河道內,裂谷、水潭、湍瀨、急流和回水都在水體和河床 中提供不同的棲息地條件。這些例子都是以物理結構來描述的,再次說明瞭結構與棲地功能之間的強大聯繫
導引功能


導引功能是作為能量、物質和生物體的流動通道的能力。河流廊道首先是由收集和運輸水和沉積物形成的管道。此外,許多其他類型的物質和生物群在整個系統中移動。
河 流廊道可以作為管道的橫向和縱向,由生物和物質在任意方向上移動。物質或動物可以進一步穿過河流廊道,從一側移動到另一側。例如,鳥類或小型哺乳動物可以 通過其植被穿過一條封閉的樹冠的河流。有機碎屑和營養物質可能從高到低的漫灘和廊道內的河流中下降,影響到河流無脊椎動物和魚類的食物供應。
移動物質是重要的,因為它影響到河流的水文、棲息地和結構,以及漫灘和高地的陸地棲息地和連接。連接和寬度的結構屬性也會影響管道功能。
對 於遷徙或高度流動的野生動物,廊道同時充當棲息地和導引。廊道與其他適當的棲息地相結合,例如,使鳴禽能夠從新熱帶的越冬棲息地遷移到北部夏季棲息地。許 多鳥類只能在休息和加油之前,在有限的距離飛行。為了使河流廊道有效地作為這些鳥類的導引,它們必須充分連接,並且足夠寬,以提供所需的遷徙棲息地。
河 流廊道也是能量運動的管道,它以多種形式出現。水流的重力驅動能量不斷地塑造和修改景觀。廊道在春夏保持涼爽,秋季更溫暖時,會改變陽光帶來的熱量和能 量。溪穀是有效的飛機場,在傍晚將涼爽的空氣從高到低海拔移動。廊道中生產效率高的植物群落作為活植物物質積累能量,以落葉或碎屑的形式大量出口。初級生 產力、營養物質流動和落葉凋落的高水準也促進了廊道的分解,從而使能源和物質發生了新的變化。在其出口處,一條河流向下一個較大水體 ( 例如,水量增加、溫度升高、沉積物、營養物質和生物 ) 的輸出部分是其自身系統的能量過剩。
最著名和研究的水生物運動和與流域相互作用的例子之一是鰻魚與鮭魚在河與海之間遷徙以產卵。鰻魚在海洋中產卵並生長呈幼體、幼魚後,溯游進入河流中游成長。 鮭魚向上游遷移產卵。在海洋中成熟後,魚類依賴於進入其上游產卵場。 就鮭魚而言,河流廊道取決於大量產卵和垂死成蟲在產卵過程中進入上游水系的生物量和營養投入。因此,連通性往往對水生物種的運輸至關重要,反過來,營養物質從海洋上游輸送到河流上游。
河流也是植物分佈和在新地區建立的管道 (Malanson 1993) 。流動的水可以在相當長的距離上運輸和沉積種子。在洪水期,成熟的植物可能被拔除、重新安置和重新安置在新的地點。野生動物還通過在廊道的不同地方攝取和運輸種子來幫助重新分配植物。
泥砂 ( 河床載或懸浮荷載 ) 也 通過河流運輸。沖積河流依賴於沉積物的持續供應和運輸,但它們的許多魚類和無脊椎動物也可能受到過多細泥砂的損害。當條件改變時,河流可能會因泥砂下降而 缺水或堵塞。缺乏適當數量泥砂的河流試圖通過下切、河岸侵蝕和河道侵蝕來重建平衡。一個結構合理的河流廊道將優化泥砂流向河流的時間和供應,以改善泥砂輸 送功能。
道路中的局部區域取決於材質從一個點到另一個點的流動。在 salmonid 示例中,產卵場附近的當地陸地區域取決於從魚類生物量向其他陸地野生動物轉移營養物質,然後進入高地。河床和水生生態系統的局部結構依賴於上游和上游的泥砂和木質物質,形成一條自我調節穩定的河道。
河流廊道寬度是重要的,因為陸地經常是大量天然泥砂和生物量的供應者。寬而連續的廊道充當一個大導引,允許沿廊道橫向和縱向流動。管道功能通常在狹窄或零散的廊道中更為有限。
篩 濾與屏障功能

河流廊道可以作為屏障,防止移動或篩檢程式,從而允許選擇性地滲透能量、物質和生物。在許多方面,整個河流廊道都是一個有益的篩檢程式或屏障,它減少了水 污染,減少了泥砂的運輸,並且常常為土地利用、植物群落和一些流動性較差的野生動物物種提供自然邊界。
進入和穿過河流廊道的物質、能量和生物體可以通過廊道的結構屬性進行過濾。影響屏障和篩檢程式功能的屬性包括連接 ( 間隙頻率 ) 和道路寬度 ( 圖 2.40) 。沿流道路邊移動的圖元在進入流道路時也可以選擇性地過濾。在這種情況下,無論是直線的還是卷積的,它都是邊緣的形狀,對濾波函數的影響最大。然而,它通常垂直於最有效地過濾或停止的河流廊道移動。
根據河流廊道的寬度和連線性,可以運輸、過濾或停止物質。物質在景觀上向大河谷移動,可被河流廊道攔截和過濾。天然植物群落結構等屬性通過吸收、吸收和中 斷,可以物理地影響流入河流系統的徑流量。廊道裡的植被可以過濾掉大部分的營養、沉積物和水的表層流動。
通 過一個河流廊道網路,可以減少較大河流中的淤積,從而過濾過多的泥砂。河流廊道過濾了許多陸地物質,使其不受阻礙地穿過景觀。地下水和地表水通過地下和地 面上方的工廠部件過濾。在河流廊道內,動植物群攔截化學元素。更寬的道路提供了更有效的過濾,連續道路在其整個長度上充當篩檢程式。
河流廊道的斷裂有時會產生漏斗破壞過程進入該區域的效果。例如,沿河流廊道的連續植被中的間隙可以通過將增加的徑流集中到該區域,從而減少過濾功能,從而導致侵蝕、溝壑以及沉積物和營養物質自由流入河流。
流廊道邊界的邊開始過濾過程。突變邊緣將初始濾波函數集中到一個狹窄的區域。漸變邊增加過濾,並將其傳播到更寬的生態梯度 ( 圖 2.41) 。
平 行於廊道的運動受不平坦廊道邊緣的凹和裂片的影響。這些功能用作流入廊道的物質的屏障或篩檢程式。單個植物在經過捲曲的邊緣時,可以選擇性地捕獲諸如風載 沉積物、碳或繁殖體等物質。例如,沿著邊界邊行走的食草動物可以停下來休息,有選擇地在一個隱蔽的角落裡覓食。風把一些種子吹進廊道,那些適合廊道條件的 種子可能發芽,建立一個人口。裂片起到選擇性過濾作用,在邊緣收集一些種子,並允 許其他物種在邊界上相互作用 ( Forman 1995) 。

圖 2.40 :植被緩衝區的寬度影響過濾和障礙功能。溶解的物質,如氮、磷和其他營養物質,進入植物體內。河流廊道受到摩擦、根系吸收、粘土和土壤有機質的限制,不能進入河道。

圖 2.41 :邊緣可以是 (a) 突然的或 (b) 漸變的。突變的邊緣,通常是由擾動引起的,往往阻礙生態系統之間的運動,並促進邊界的運動。漸變邊緣通常出現在自然環境中,更加多樣化,並鼓勵生態系統之間的運動。
來源與匯聚功能
