Streams and Life 河流生物與生態
www.epa.url.tw 林雨莊編 著
B01 河流的物理環境
B02 非生物因素
B03 河流裡的能源
B04 河流食物網
B05 河流生態結構與功能
B06 河流類型與環境
B07 淡水藻類與微生物
B08 大型水生植物
B09 水生昆蟲
B10 河流軟體動物
B11 河流甲殼動物
B12 爬蟲和兩棲類
B13 河流魚類
B14 河流鳥類
B15 河流哺乳類
B16 河流的生態威脅
第二章 非生物因素
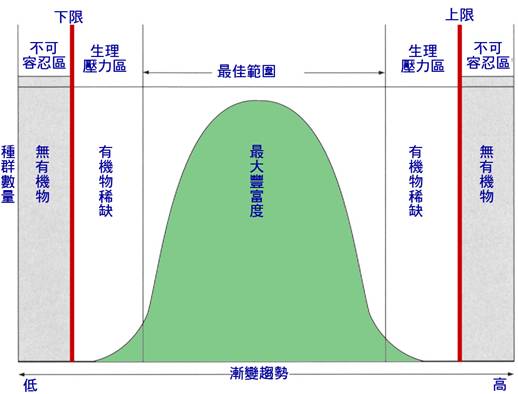
圖 2.1 在某些非生物變數 ( 如溫度、流速等 ) 的條件範圍內,某一特定物種個體表現的概念模型。如果因數是溫度,則冷適應物種的最優值將向左移動,暖適應物種的最優值將向右移動。
生命是否繁盛或稀少有限,取決於物質環境。正如河流在大小、坡度、流量和河道特徵上表現出巨大的物理變化一樣,河流在物理和化學因素的大變化,統稱為非生物因素。這些因素包括光合作用所必需的陽光、水流、溶氧,它可能把食物帶給魚類,或使昆蟲從河床上掃到魚類的視線中;溫度,對活動水準和生長速度至關重要;河床基質,它創造特定的棲地;以及各種化學因素,從營養到污染物。可能的組合是無限的,為每一條河流提供了獨特性,並對生態工作者在尋找模式和普遍性方面提出了巨大的挑戰。在這一章中,我們將繼續探索主要的非生物因素,結合水文和地貌變數,為河流生態創建物理範本,從而為河流中的生命發展物理環境。
圖 2.1 顯示了一個考慮非生物因素的有用的,儘管是簡化的模型。這個模型設想了粒子的最佳範圍及變數 ( 這也取決於特定的物種,因為生物體可能在生理上適應,在不同的因數範圍內表現最佳,就像高海拔冷水魚和低海拔溫水魚一樣 ) 。在最佳範圍之外,生物體會遭遇到水溫、鹽度、酸鹼度、溶氧度等可容忍範圍,不可容忍的壓力增加,最終導致致命。這個模型在很多方面過於簡單,曲線的中間部分可能比鐘形部分更平坦,曲線可能不對稱,自然界中的生物必須同時處理許多變數。如果任何一個因素是極端的,我們可能會發現很少有物種能夠忍受環境條件;多樣性和生產力將很低。我們可能會遇上景觀美麗的高山湖泊或溪流,但可能沒有生命,可能是碳酸鈣或某種化學物質濃度、溫泉或污染的酸雨所造成的。在大多數自然生態系統中,沒有一個因素是永久性限制的,可能雨水沖淡或化學濃度降低而改變。我們更經常看到各種非生物因素重要性的季節和空間變化。冬季溫度可能限制活動,早春光照可能充足,但在樹冠層葉片脫落後的環境條件改變,等等。當所有非生物因素都在大多數物種的可容忍範圍內時,可能的因素組合的虛擬無限可能有利於物種的不同組合,影響生產力,並有助於解釋使每條河流在某種程度上具有獨特性的奇妙生態變化。
2.1 陽光
在所有的非生物因素中,光是最容易描述的。這是光合作用所必需的。光也可能是一個棲息地的因素。魚類或無脊椎動物可能會避開溪流中陽光充足的地方,也許是因為它們更容易被捕食者看到。有時,蜉蝣若蟲聚集在陽光充足的地方,可能以大量的藻類為食。但光對植物生長的影響是其主要作用,洞穴溪流很好地證明瞭這一點。在一個無休止的黑暗洞穴中,生命依靠極少量進口的有機碎屑及其相關微生物生存;水生植物幾乎不存在。人們只能在洞穴溪流發現非常低的物種密度,通常是撓足類或水蚤。洞穴溪流當然是一個極端,但是很少日照的溪流是很常見的,源流或上游段的溪流很在森林茂密的山區。我們可能不經常在河流上游茂密的森林樹冠下步行或釣魚。河道因為遮蔭,光線很難透入,水體中植物、藻類也就非常稀少,當然也造成水棲生物及魚類的稀少。
當然,光合作用所需的光照是隨季節而變化的,並且受任何可能阻止光照到達河面和河床的因素的影響很大。森林景觀中的小河被森林本身或被狹窄山谷的峭壁遮蔽。如果森林是落葉性的,藻類可能在冬、春季生長旺盛,而在夏季樹葉茂盛時生長較少,也許在秋天落葉後又會短暫生長。如果森林是茂密的針葉林,,那麼小溪在任何時候都可能非常暗。僅有少數的藻類 ( 矽藻等 ) ,仍然能夠適應微光。
在相當陰暗的條件下有些增長。正如我們在第 3 章中更詳細地描述的那樣,秋季落葉和其他非生物有機物來源常常取代光合作用,成為這些高度遮蔽的溪流中水生食物網的能量基礎。在許多小溪流中,光限制是一個可變因素,在某些地方或季節比在其他地方或季節具有更大的意義,並且部分地被一些藻類的適應抵消,以允許在弱光下進行光合作用。
較寬大的溪流往往較少被森林和山谷的牆壁遮蔽。在寬度為 10 或 20 米的地方,一天中至少有一部分時間會有大量的陽光照射到河面。這些環境有利於很高的光合速率 ( 例如桶後溪、大豹溪、清水溪、內雙溪等自然溪流上游段,水質良好卻呈現綠色,河床塊石長滿附著藻類 ) 。但隨著河流的加深,光線必須穿透更深的水域才能到達河床。水體中的懸浮物濁度使光衰減;在非常渾濁的河流中,這種情況更可能發生 ( 例如淡水河華江以下的主流河道,河床缺乏可供藻類附著的石塊,陽光透入到河底的光照量也很少 ) ,藻類和沉水有根植物都不會生長良好,除非在淺水區,甚至浮游生物 ( 懸浮藻類 ) 也會嚴重不利。某些河流段可能相當湍急,因此這些浮游生物不斷地被捲進陰影深處。它們可能不時地被掃進光線充足的淺水中,短暫地進行光合作用,也可能是浮游生物在呼吸空氣後,然後再次被掃進黑暗中。
2.2 水流
水流是溪流的物理變數。它似乎與圖 2.1 的模型很好地吻合;許多生物體佔據一定的水流速度範圍,並且似乎受到極值的壓力。但水流是一個複雜的物理變數,因為它與許多其他非生物和生物因素相互作用。生命有機體 ( 魚類、水生昆蟲 ) 必須躲避或抵抗水流,開發附著裝置,或執行耗體力、耗熱能的工作來保持其位置。水的流動將氧氣輸送到有機體,這可能有助於它們的呼吸。同樣的流量將二氧化碳和營養物質輸送到植物體內,刷新了它們對光合作用的需求。正如第 6 章所描述的,每隻魚都知道,水流將食物輸送給等待的有機體,比如躲在岩石後面的魚類,當一個足夠誘人的食物漂來時,它就準備捕捉。水流速度也塑造了河床棲息地,侵蝕了這裡的河床基底並沉積在那裡,形成了礫石淺灘、樹幹淺灘 ( 湍瀨 ) 、平流帶和深潭。水流的速度決定了細顆粒是否會沉降到河床面上,從而導致一種棲息地退化,即淤泥填充空隙並覆蓋表面,雖然河底的細砂或淤泥也能生長附著藻類,但是因基質不理想、易受擾動,附著藻類生長量要少很多。水流的重要性,加上其影響的複雜性,使得人們對這一關鍵的河流變數進行了大量的研究。
它看起來很簡單,可以測量水流,然後研究其影響。不幸的是,我們對水流運動的研究越多,這個課題就越具有挑戰性。當雨水或融雪向河道輸送更多的水時,水流速度會增加,這可能是季節性的規律性或偶然性的不可預測性。水流在河流彎曲的內側和外側流速大不相同,在湍瀨和深潭之間,以及每一塊倒伏木、岩石或其他障礙物的前、後、邊側都不相同。有岩塊、樹枝堆積在河床的生態非常多樣,在那裡幾乎所有的藻類、無脊椎動物以及許多魚類都能被發現。最令人困惑的是河床表面或各種障礙物表面的流速,但是它們非常難以測量,因為很難找到量測 mm 等級水流量測儀器。此外,我們從使用染料和最先進精準的測量中知道,精細尺度的水流速度是難以置信的非均勻性。理論告訴我們,與石頭或其他固定物體表面接觸的水分子層不會移動,而些微上層的水分子層會移動;移動的和不移動的分子之間的摩擦會產生湍流和剪切立,從而影響生物體。 mm 毫米級的流量測量只能在精心控制的實驗室環境中進行,而沒有大小和紋理不同的石頭所帶來的結構複雜性。但是,目前正在取得進展,生物體的適應能力告訴了我們許多當前影響它們生活的方式。
居住在溪流中的動物表現出各種各樣的適應能力,以抵抗水流,並可能利用它來獲得優勢。無脊椎動物身上有各種各樣的附著裝置,包括石蠶蛾吐絲、螺類、蝸牛的吸盤和其他粘性分泌物、鉤、爪等。平滑流線型的體型也可以使生物體避免一些剪切應力,儘管這是不確定的。流石蠶幼蟲 ( 圖 2.2) 能忍受很高的水流速度。通常它們在快速、平滑的水流,甚至在垂直面上。這些幼蟲附著在後半身上,後半身上有一圈向外的鉤狀物,這些鉤狀物嵌在一層絲中,然後旋轉到石頭表面。如果有水生昆蟲被水沖移動,一條絲線可以讓動物重新拉回獲得附著的表面。黑蠅幼蟲從水中獲取食物,它們用附著在頭上的扇子過濾剛毛,稱為頭扇子。一般來說,通過梳扇的水越多,過濾的食物就越多,因此生活在快速水流中的能力提供了真正的好處。
魚類在快速流動的水中表現出各種各樣的適應能力。快速遊動的魚類和在快速水流中遊動的魚類的橫截面呈流線型和橢圓形,或者像爬岩鰍用腹部貼附在急流的石頭上 ( 圖 2.3a) 。它們與身體較深的翻車魚 (sun fish) 形成鮮明對比,翻車魚 ( 曼波魚 ) 生長在較慢的河流、水潭塘和湖泊中,比速度更適合機動。許多生活在河底的魚類,利用其粗糙度作為庇護所,表現出一套特殊的適應能力。許多人的膀胱縮小或沒有,失去了浮力。身體在背腹平面是扁平的,眼睛是背向的,胸鰭經常是擴大的,肌肉發達,以幫助保持位置對抗水流 ( 圖 2.3b) 。在實驗室水槽中的實驗表明,隨著水流速度的增加,這種魚可能會拱起背部,並利用其肌肉胸肌,以防止滑脫。鯰魚和其他種類的魚都具有這種適應能力。附著吸盤存在於許多脊椎動物中,包括鰕虎科和兩棲蝌蚪。
這些例子說明,魚類和無脊椎動物都有很多方法來抵禦水流。兩者都能熟練地避開水流。魚類被證明有利於在靠近高流速的慢水流中覓食 ( 用魚類生物學家的話說,它們在水中“站住” ) 。它們在保持位置上消耗很少的能量,同時使通過它們視野的食物顆粒數量最大化。曾經有人認為,在石頭表面附近存在的一個小水流區域允許一個具有扁平輪廓的無脊椎動物來避免水流力。現在看來這似乎不太可能,但扁平的形狀肯定能讓昆蟲在石頭下面和石頭之間移動,從而避免一些水流。
水流對藻類和微生物有多大影響?由於它們體積小,給實驗研究帶來了更大的困難。我們知道洪水會破壞附著的苔蘚 ( 滾石可能倒地,附著藻類會因洪水擾動懸浮泥砂覆蓋石塊,附著藻類因缺乏陽光而死亡;因此,“滾石不會聚集苔蘚” ) 和藻類,它們在高流量事件中會被沖刷,並從基質中脫落。但是流動的水提供了新鮮的營養物質,並去除了有害的代謝副產物,所以適度的水流速度可能是有益的。在實驗室水槽和一些穩定的自然系統中進行的研究,記錄了藻類墊席的持續堆積,上層生長和分裂的細胞附著在光照和營養缺乏的衰老細胞的底層。隨著生物墊變厚,它的質量增加,死細胞層變大,最後生物墊 ( 藻類墊席 ) 脫落。稍快的水流很可能將營養物質更新到更深層。然而,更強的水流會導致它脫落。
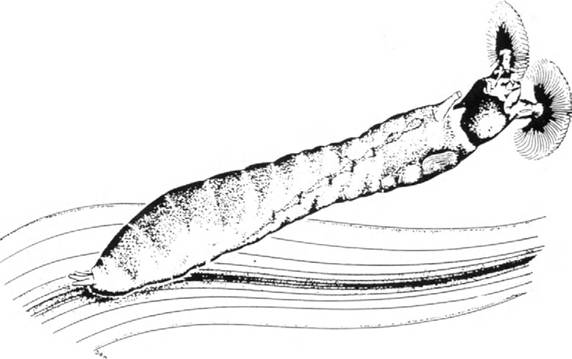
圖 2.2 黑蠅幼蟲對急流中生活的適應。在其典型的過濾姿態下,幼蟲被嵌入絲中的鉤子所附,被水流強迫接近水準。它的身體有點扭曲,可以通過沿著腹神經索的線看到。頭部的過濾風扇捕捉到像細菌一樣小的顆粒。

圖 2.3 (a) 適於在水體中逆流游泳的流線型魚類
(b) 一種底棲魚,通過其擴大的胸鰭適應於靠在基質上的位置。
表 2.1 石塊或卵礫石大小分類
石塊名稱 顆粒大小 (mm)
大岩塊 Boulder >256
大卵石 cobble 128-256
小 鹅 卵石 64-128
大卵石 pebble 32-64
小卵石 16-32
粗礫石 grave 8-16
中礫石 4-8
細礫石 2-4
極粗砂 sand 1-2
粗砂 0. 5-1
中砂 0. 25-0.5
細砂 0.125-0.25
極細砂 0.063-0.125
淤泥 / 黏土 silt <0.063
2.3 河床基底
岩石、砂礫、泥土、樹木、塑膠物都可能是構成河床的基質,藻類、微小動物附著生長,建構河流食物來源的第一層次。河流的無機基質是由水流輸送、分類和沉積的地質母質決定的。樹幹、枝葉,起源於河岸和河灘地、運輸、沉積,也許部分掩埋,是主要的有機基質。所有這些河床成分都是潛在的障礙物,它們會產生不均勻的水流,並充當藻類和微生物生長的附著表面。仔細觀察河床,或者涉水走一段距離,用腳感知河床。石頭的大小、形狀和粗糙度的差異對我們來說是顯而易見的;想像一下這對水生昆蟲構成了什麼樣的棲息地。那塊石頭傷了你的腳趾,對一些魚來說是個很好的庇護所。落水的植物,從樹枝到樹幹,在許多溪流中都很豐富,儘管自然的河岸森林可能不存在或已被清除。幾根小樹枝,加上一些秋天的落葉和一些細砂,就可以為石蠅和蠅、蚊幼蟲創造一個很好的小棲息地。一棵倒下的樹和它的根莖可以在一條大的河流岸側中形成一個深潭,可能容納一些魚棲息,使一些垂釣者感興趣。
無機粒子的基質可以很容易地根據粒子大小分類。通常使用的地質比例尺 ( 表 2.1) 可以量化基底樣品 ( 並告知我們小卵石比粗礫石大 ) 。巨石更多的是山溪的一個特徵,在那裡它們與陡峭的坡度結合形成階梯水潭 ( 圖 11.6) 。鵝卵石和礫石可能是三至四級序溪流的典型,而細砂通常佔據著低地平原河流 ( 五級序以上 ) 的河床。
許多河流的平緩河段是細砂底的,而且細砂是可用的建築材料。淤泥質和粉質底質是典型的非常緩慢的水流,因此它們最常見於回水和側溝。相反的,粘土質可能形成平滑的河床,如果地質條件好的話,河床會很湍急。這是因為粘土是粘性的,儘管它們的顆粒體積很小,在急流中形成一個密實而且非常光滑的河床表面。
深潭和湍瀨進一步證明,水流通過日流量以及沖刷和沉積循環影響河床物質。在低流量期間,較細的物質在水潭中沉澱,因為水流的速度低於微粒沉降的速度。在河灘中,水流足夠迅速,以防止沉降,並收集 ( 夾帶 ) 了大多數砂礫物質。
流動的水總是會輸送一些無機物質,從最清澈的山澗到混濁富含泥砂的河流。生物體可能表現出對特定底物的適應性,這在大型無脊椎動物中最為明顯。某些物種與附著藻類、細砂、鵝卵石混合物或大塊石有關。許多研究試圖更精細地測量河流生物對基質的偏好,可參考表 2.1 。一些物種對快速水流中的大鵝卵石表現出一定的親和力,但通常這種聯繫最好用統計趨勢來表示。對於許多大型無脊椎動物來說,細砂似乎是一種貧瘠的基質,儘管有一些特殊生物,如穴居的蜉蝣、顫蚯 ( 紅蟲 ) ,和許多非常小的,主要是間質分類群會生活在細砂或淤泥質的河床中。但是與岩塊基質、卵石基質相比,砂質基質的多樣性和生產力普遍降低。它的不穩定性使細砂成為無脊椎動物生存和豐富食物供應的貧瘠之地。譬如宜蘭冬山河河道治理後,河道管道化,河床清除了岩塊與雜物,平滑的河床,使許多本土魚類失去食物來源,外來種琵琶鼠、吳郭魚等充斥中下游河段。
河流中的任何基質都可以提供藻類生長的表面,而河床的任何粗糙度都可以提供有機物和微生物可以在其中積聚的沉積袋。因此,基質對溪流食物網的營養至關重要。穩定的表面通常支持較大的藻類生物量。河床的粗糙度在當地條件下造成了異質性,從而增強了多樣性。
最後,必須認識到倒伏木作為基底和通道結構決定因素的重要性。很難說倒伏木對不同種類的河流有多重要。在許多地方,河岸森林的砍伐和洪水的控制已經減少了河流中大量植物碎片的供應。但是許多證據表明,植物樹幹枝葉是河流中有機物的重要來源。砂質河底附著藻類很少,相對來說是很少生產力的;這些系統中的無脊椎動物生產與障礙物密切相關,魚類也是如此。在低人為幹擾的自然河流中,木材在河道中更為豐富,當時它很可能創造了棲息地,塑造了河道,並充當各種生物的基質。台灣大多數較大溪流都在河流出山口建造了水庫,意味著水庫裡聚積了大量的倒流木、樹幹枝葉,水庫下游至出海口河段很難出現大型倒流木,對於水生物棲息環境有較不利的影響。
2.4 水溫
水流溫度很重要,原因有很多。由於很少有淡水生物是溫血的,水溫對幾乎所有生物的代謝率和整個系統的生產力都有很大的影響。生理適應使魚類能夠在相對較低的水流溫度下活動和生長,而平頷鱲等魚類在稍高的溫度下表現最佳。在寒冷的溪流中,蜉蝣通常以滯育胚胎的形式越冬,每年完成一代,而在溫暖的溪流中,卵期較短,若蟲全年活動頻繁,一年中可能發生好幾代。因此,在下個世紀預測的氣候變暖可能會改變河溪的生態,因為暖適應物種取代冷適應物種;系統生產力也可能也會增加。
在未來氣候變暖的情況下,人們有理由擔心失去適應寒冷的物種。一些物種肯定會向北遷移,但集水區分水嶺邊界和東西河流可能會限制它們的機會。
熱帶溪流的河水溫度可能達到 30 攝氏度,亞熱帶大多數溪流的年溫度範圍在 15 到 25 攝氏度之間。生物能夠生存的水溫上限取決於它們的生活史和溫度適應。 魚不是恆溫動物,它的體溫隨著水溫變化而變化,與水溫的溫差不大於 1 度。 高身鯝魚、櫻花鉤吻鮭偏好低於 20 度的水溫 ;粗首鱲 最適宜水溫 20-25 度;鯽魚:最適宜水溫 16-25 度;鯉魚、鯰魚:最適宜水溫 20-30 度。超過最適宜溫度上下限 5 度,則容易停止攝食或死亡。 大多數溫水魚的上限接近 30 ℃,非洲來的熱帶吳郭魚可以耐受 25 至 35 ℃。 總的來說,河流淡水魚在 15-25 度的水溫比較適宜。魚會隨著水溫度的變化而調節自身體溫,有兩種調節方式: (1) 生理調節:水溫發生變化時,魚的敏感器官利用身體體液,調節體溫儘量與周圍水穩定一接近。 (2) 動態調節:因為水溫與氣溫有差異,水的密度比空氣大,所以散熱比氣溫慢,加上水分上、中、下三個層次,這三個層次溫度也有差異,水越深,差異就越大。據數據測量,當水的上層溫度在 21.2 度時, 1 米深的水溫為 18.3 度, 2 米深的水溫為 17 度。在冬季,水的上層溫度為 4.3 度時, 1 米水深的溫度為 4.5 度, 2 米水深的溫度為 4.9 度。當水溫不適宜某種魚生存時,會尋找自己適合的水層。。
水溫首先受氣溫的影響,然後受地表水和地下水的混合、遮蔭程度和溪流大小的影響。高緯度或海拔更高的河流當然更涼爽,因為它們的氣溫和太陽輻射更少。地下水的溫度通常非常接近年平均氣溫,因此春天的小溪溫度非常恒定,夏季涼爽,冬季溫暖。隨著地表和淺層地下水流成為河河流文收支的一個較大部分,它開始呈現月平均氣溫的季節特徵。由於河床的太陽能加熱和河流中相對較小的水品質,小溪流白天變熱,晚上變冷,造成晝夜溫度變化大,這種變化可以相當明顯。水的熱慣性比空氣大得多,所以它經歷的劇烈“擺動”較少。當許多小溪流匯成大河流時,水的品質慣性對這種變化起緩衝作用,因此晝夜變化最終變得非常小 ( 圖 2.4) 。最後,由於太陽輻射能到達河流表面和河床,遮蔭的河流確實更涼爽,沒有河岸植被的河流確實更高溫。幾公里的樹蔭足以產生明顯的差異,並決定魚類是否能佔據該河段。魚類會根據非常小的溫差來選擇微棲地,這大概是因為它們在當前的環境中稍微降低了它們的代謝成本的生理優勢。
2.5 化學因素
河流的化學成分首先取決於其集水區的地質情況,但雨水和人類活動也可以提供重要的輸入。有很大的變化,特別是在小溪流的上游,土壤地質化學物質溶入,影響較大。以下我們將描述溶解氧、鹼度、營養素和人類污染物。
氧氣是大多數生物體的關鍵需求,通常不是一個限制因素。它在大氣中含量豐富,易溶於水,只有靜止且深的水體,溶氧不能在水的表層和深層之間循環,且水中生物消耗量大時,才會顯得不足。小而湍急的水流通常處於飽和狀態。如果代謝活性很高的話,大而流暢的河流可能會在底部附近經歷一些缺氧。飽和值隨著水溫的升高而降低,在高海拔地區,大氣氧分壓小於低海拔地區,飽和值降低。很高水準的有機輸入,如果河水中含有人類或動物糞便的污水,或加工食品的廢棄物,會產生高量的細菌代謝,導致於所有的氧氣都被消耗殆盡,就會產生腐敗的臭味。
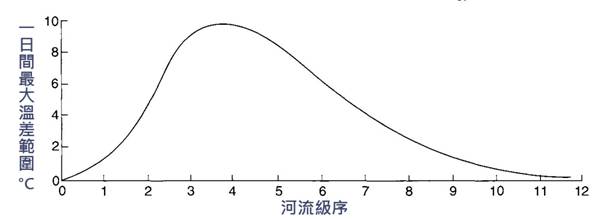
圖 2.4 溫帶河流日最高氣溫與河流順序的關係。
酸鹼值是一種測量化合物的種類和數量的方法,顯示 氫離子濃度指數 , pH 值為 7 是中性的,而較低的值是酸性的,較高的值是鹼性的。如果一條溪流落葉與水中植物特別多,過多的植物物質的分解就會呈現褐黑色的水質,或稱為黑水溪是天然的酸性河流,當接觸酸雨或一些採礦作業產生的碎石時,溪流也會變成酸性。肥沃土壤的排出來的溪流通常是微鹼性的,流經白堊土壤的溪流可能是強鹼性的。鹼度主要受水中碳酸氫鹽和碳酸鹽含量的控制,這就解釋了為什麼類如廣西桂林的石灰岩 ( 碳酸鈣 ) 河流顯示出高鹼度。一般來說,鹼性水越多,生物生產力就越高,酸性水越多,生物生產力就越低。在一些受酸雨影響較大的地區,為提高湖泊的生產力和中和湖泊的酸性,通常採用石灰法。它偶爾也會在河流中使用,但是在連續流動的水體中不太適用。
營養素是某些有機體生命所必需的元素,尤其是植物和微生物在生長過程中,產生的生物量將支持食物網。維持生命所必需的元素和維生素的清單相當長;通常我們主要關注氮和磷,它們以能使湖泊、水庫變綠而聞名。
實驗室水槽試驗表明,營養鹽濃度的變化會影響藻類的種類組成和總生物量。當光線充足且水流不太快時,密集的絲狀藻類的長羽狀藻類表明營養過剩。事實上,許多溪流和河流似乎攜帶了足夠的氮和磷,以刺激大量甚至過度的藻類生長。在岩石相低溶解的小溪,可能營養素濃度較低,不足以支持藻類或水生植物生長,也不適宜作為農業灌溉水。河流中的藻類可能受到許多因素的影響,如陰影、飛濺、基質移動、溫度或禽畜養殖,我們往往不會看到河流優養化程度達到湖泊中的程度。但是當河流進入水庫,水流緩慢,允許沉積物沉澱時,密集的藻華告訴我們,水含有大量的營養物質。這些營養物質的影響可能是非常嚴重的,因為在藻華過後會死亡、下沉和腐爛。其結果可能是由於細菌代謝而在深水中形成缺氧區,與大量污水排入湖泊或海洋的情況沒有什麼不同。
今天,人類活動對河流的營養負荷有很大貢獻。磷主要來自污水輸入和土壤侵蝕。廢水處理廠已經有效地減少了磷的輸入。改進的農業和“最佳管理措施 BMP ”做法在減少土壤侵蝕方面取得了一些成功,土壤侵蝕會使磷附著在土壤顆粒上。這突出了點源污染,如污水,在技術可以集中的地方進入河流,而非點源污染,從多個擴散源進入河流,因此需要許多個人的共同努力來控制它。目前,非點源污染仍是河流養分負荷的主要貢獻者。
氮也是一種重要的營養素,儘管在一段時間內它比磷受到的關注要少。作為一個粗略的概括,有證據表明磷的富集導致湖泊優養化,而氮的富集對海洋水域有相應的影響。美國有《清潔水法》,台灣有《水污染防治法》約束水質排放,但是密集人口的都市地區或工廠區容易建造污水收集系統及污水處理廠,但是廣大的鄉村或高山散置農村等非點源污染,防治難度較大。並且,氮很難控制。通過製造肥料和燃燒化石燃料 ( 產生氮氧化物 ) ,人類現在控制著世界的氮流量。此外,還有非常大的大氣輸入,來自化石燃料工廠、汽車尾氣等。在風和大河的推動下,交換發生在非常大的範圍內,同時我們所有的汽車中的氮氧化物也被雨水輸送沉積。
化學賀爾蒙 ( 激素 ) 是河流中最後一類不愉快的非生物因素。與營養素一樣,有些是非點源,而另一些是點源。酸雨可能降低溪流的 pH 值 ( 增加了酸度 ) 。一部分是點源 ( 二氧化硫和燃燒化石燃料的發電廠產生的氮氧化物 ) ,另一部分是非點源 ( 汽車尾氣產生的氮氧化物 ) ,通過雨水傳播。有些土壤有足夠的鹼度來防止其產生很大的影響,但當酸雨落在堅硬的火成岩集水區時,河水的化學特徵是酸性的,就像雨水一樣。河流中的生物環境也是貧瘠的。
採礦是酸性和有毒金屬的另一個來源,它們一起毒害了礦區下游的許多溪流。礦石開採後留下的碎石易被雨水風化。這會在溶液中產生酸和金屬,這些酸和金屬會產生劇毒,甚至在河流的大部分區域幾乎都會導致生命滅絕。
南臺灣的養豬事業興盛,豬的糞尿排放量是人的 4 倍,數百萬隻豬使許多溪流發黑、發臭、豬糞塊漂浮於河面。較大規模的養豬戶可以輔導建造污水處理,但是七成以上的養豬戶都是小規模養殖。由於高屏攔河堰為飲用水源,高屏溪中上游有離牧輔導與較嚴格的管控,其餘溪流水質普遍不佳。數千隻的養鴨、養雞、養魚場也是河流的重大污染威脅。我們可以繼續從家禽和家畜養殖場、農業、建築業的廢水和化學污染物清單,以及各種各樣的工業活動,無論大小。我們對污染的認識提高了,更嚴格的法律也制定了,沒有人願意生活在被污染的溪流或河流旁。科學是明確的,一些法律和政策工具已經實施。對於河流環境保護意識還需要再加強,執法面也要堅持不懈。