
魚類生態學 (2)
1. 水環境與魚分類
2. 魚類生理學
2.1 魚的分類
2.2 魚的生理功能
2.2.1 魚類的形體
2.2.2 魚類呼吸系統
2.2.3 魚類血液循環系統
2.2.4 魚類消化系統
2.2.5 魚類排泄系統
2.2.6 魚類神經系統
2.2.7 魚類感覺器官
2.2.8 魚類生殖系統
2.3 魚類外部器官
椎、頭、顎、眼、鰓、皮、
鱗、 側腺、螢 斑、鰭、鰭線
2.4 魚鰭的類型
2.5 魚類內部器官
腸、幽門、胃、腎、脾、
肝、心、膘、韋伯
2.6 魚的疾病與免疫
2.6.1 疾病與寄生蟲
3. 環境條件對魚種分佈的影響
4. 生物因數與魚類群落結構
5. 移棲地 與魚群行動
6. 攝食與生長
7. 生活史和種群動態
8. 魚類應用生態學
第二章 魚類的生理
最早可歸類為魚類的生物是在寒武紀期間首次出現的軟脊索。儘管他們沒有真正的脊椎,但他們擁有脊索動物,這使它們比無脊椎動物更加敏捷。魚類將在整個古生代時代繼續發展,並多樣化為各種各樣的形式。古生代的許多魚類都發展了外來盔甲,以保護它們 免受掠食 者侵害。第一批 具顎的魚 出現在志留紀時期,此後,許多 ( 例如鯊魚 ) 變成了強大的海洋掠食者。
大多數魚 是放熱的 ( 冷血的 ) ,它們允許體溫隨環境溫度而變化,儘管某些大型 活躍的 游泳者 ( 如 白鯊和鮪魚 ) 可以保持較高的核心體溫。
魚類可以通過聲音在水下環境中進行通信。魚體中的聲音交流將聲音 信號從一個個體傳輸到另一個體。聲音的產生是魚類之間交流的一種手段,最常用於餵養、侵略或求偶行為。魚發出的聲音會根據所接觸的物種和刺激而變化。他們 可以通過移動骨骼組件產生聲音,或者可以通過操縱專門的器官,例如非發聲器的聲音 鰾 。
魚類在大多數水域中都很豐富。在幾乎所有的水生環境中都可以找到它們,從高山溪流的鱒魚,到最深的海溝 ( 例如,海鰻和安康魚 ) 。在已知的 34,300 種物種當中,魚類比其他任何脊椎動物群體都具有更大的物種多樣性。
魚類是全世界人類的重要資源,尤其是作為食物、蛋白質。商業和自給型漁民在野生漁業中捕撈魚類,或將其養殖在池塘或海洋 的網箱養殖 。部分魚種也扮演休閒、寵物娛樂、水族館、科學、教育的角色。
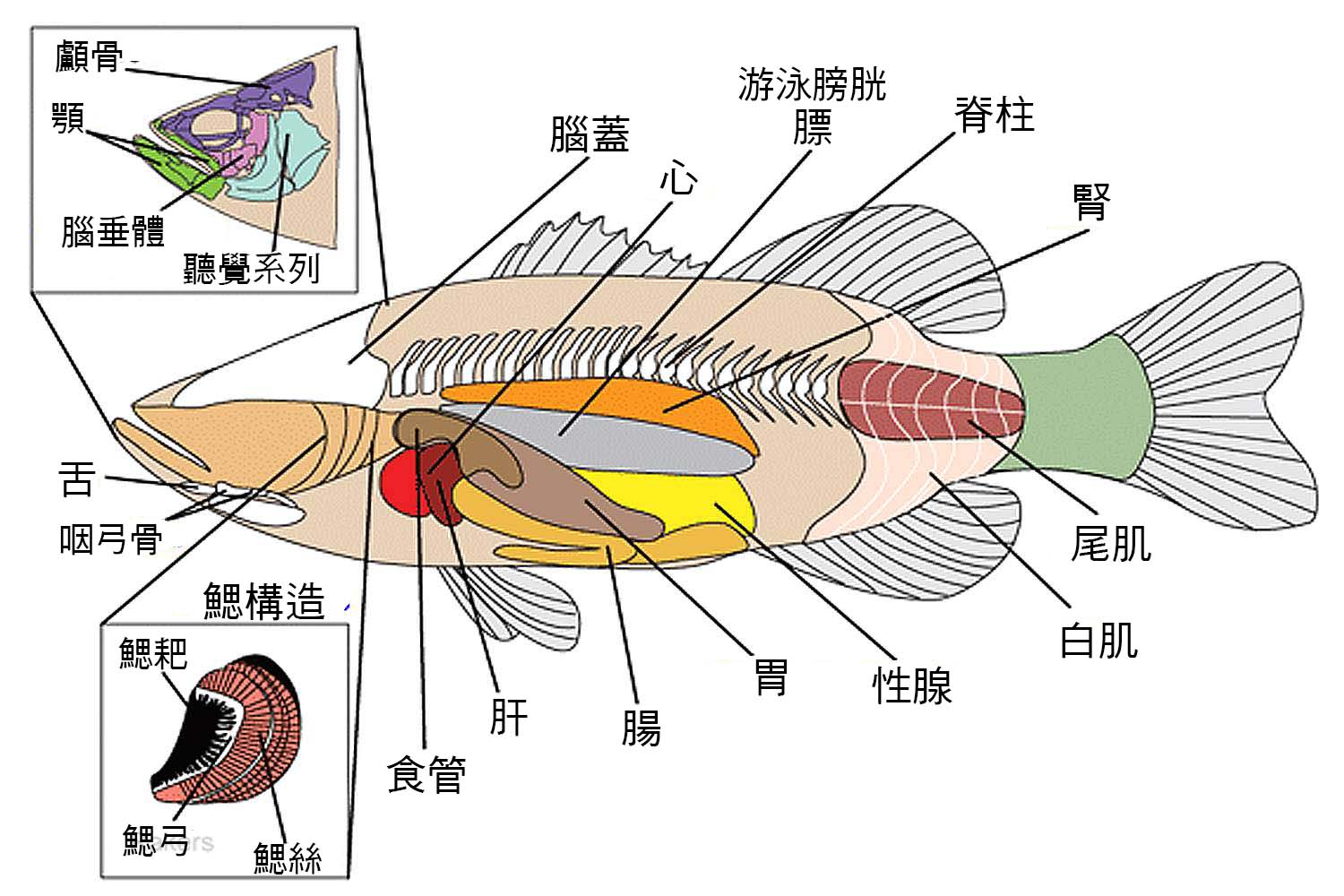
圖 2.1 魚的解剖圖
2.1 魚的分類
魚是 共生類群 :也就是說,任何包含所有魚的 進化枝 也都包含 陸地脊椎動物 , 在科學的分類法中,例如 人類 、 恐龍 等也是屬於 硬骨魚 演化而分支 。傳統的分類將魚類分為三個現存的類別,並且某些物種的滅絕形式有時在樹中進行分類,有時也將其歸為自己的類別:
(1) 無顎魚類 ( Agnatha 類 ) ,如七 鰓鰻 、
(2) 軟骨魚類 ( Chondrichthyes ) ,如鯊魚、 鰩 魚 ( 魟 魚 ) 、 肺魚 ( 原始的肉鰭魚 )
(3) 硬骨魚類 ( Osteichthyes ) ,如 腔 棘 魚、鯉魚等 。
地球上的脊椎動物可概分為魚類 (53%) 、鳥類 (18%) 、爬行類 (12%) 、哺乳類 (9%) 、 兩生類 (8%) 五大類。目前全球已命名的 魚種約在 32100 種。地球上各種魚類占脊椎動物種類的一半以上。 已知近 28,000 種現存物種,其中近 27,000 種為硬骨魚類,其中有 970 條鯊魚,魟魚等,以及約 108 種無顎魚類。 硬骨魚類最多,並且有 三分之一屬於九個最大的科;從最大到最小, 這些科 依序是鯉科、 鰕虎 科 、 慈鯛科 、 脂鯉科 、 甲鯰科 、 爬鰍科 、 鮨科 、 隆頭魚科和 鮋 科。大約有 64 個科僅包含一種。現有物種的最終總數可能增長到超過 32,500 個。
2.2 魚的生理功能
2.2.1 魚類的形體
魚類共通的特徵是有 鰓 的 水生動物 ,缺乏四 肢 及末端的 指 頭,且皮膚表層含有細毛。 魚的形狀各種 各樣, 有時相差大,但總的來說 大多數魚呈細長 的流線形狀,一般在水中速 游 的魚身體細長,而慢 游 或在水底生活的魚比較扁平。但也有的魚的形狀非常出奇,比如 海馬 。最小的魚不到 1 厘米左右,最大的 鯨鯊 可以達 18 米。魚的尾鰭是控制魚的速度與平衡的器官,其形狀更是左右上述功能的所在,一般而言,剪刀狀的尾鰭, 游 速度最迅速,如:鮪魚 及 劍魚 、旗魚,因為剪刀狀的尾鰭面積較小,與水摩擦也相形變小,反之像 孔雀魚 等,尾鰭太過龐大,因而造成其 游 速變慢。
2.2.1.1 魚的骨骼
魚的骨骼是由 軟骨 ( 軟骨魚 ) 或硬骨 ( 硬骨魚 ) 構成的,在頭骨的兩邊有四至七片 鰓 ,其中最前面的一片演化成了下 劾 骨。魚的脊椎骨是與頭骨連在一起的,在胸部有肋骨與脊椎相連,在背部,尾部和腹部有從脊椎伸出的長的刺。硬骨魚在肌肉內常有硬化的 脛 所構成的魚刺。魚 使用 鰭 來控制 它的方位和運動。大多數魚 的鰭內有 骨質 的刺來加強 。胸鰭和腹鰭是成對的,並通過肩和 髖的 肌肉相連。背、尾和 肛鰭與 脊椎相連。有些魚背和 尾之間的鰭內沒有刺,但可以硬化成角質以得以加強。魚運動時主要依靠身體的擺動和尾鰭。
2.2.1.1 魚的皮膚
魚有兩層皮膚,表層的皮膚內含有能夠分泌粘液的腺,內層有許多連接組織, 鱗和 色素細胞 也在這一層里。外層的粘液幫助魚減輕其游泳時的阻力。軟骨魚沒有粘液,但它們皮膚上細小的、牙齒般的突起有類似的作用。
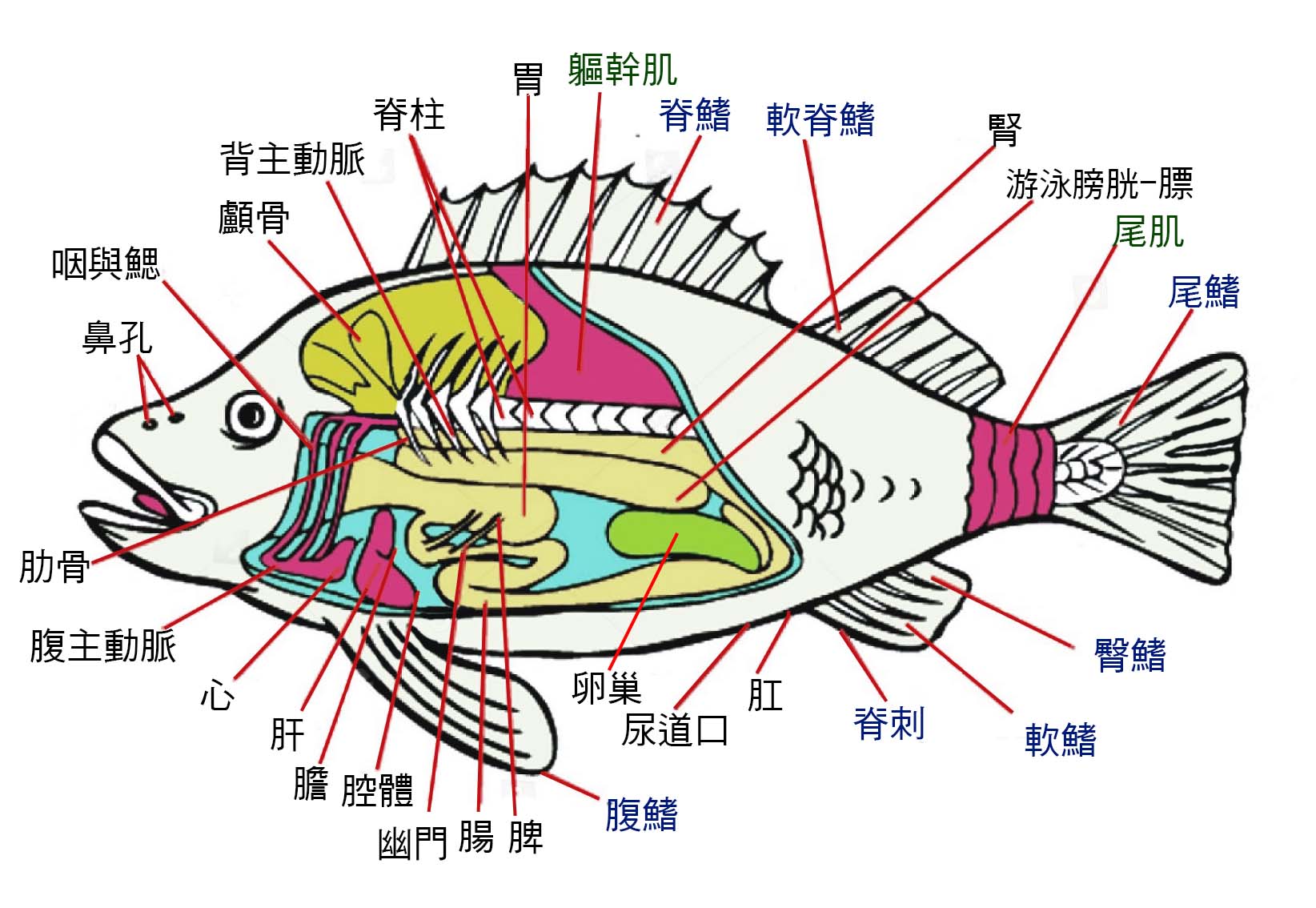
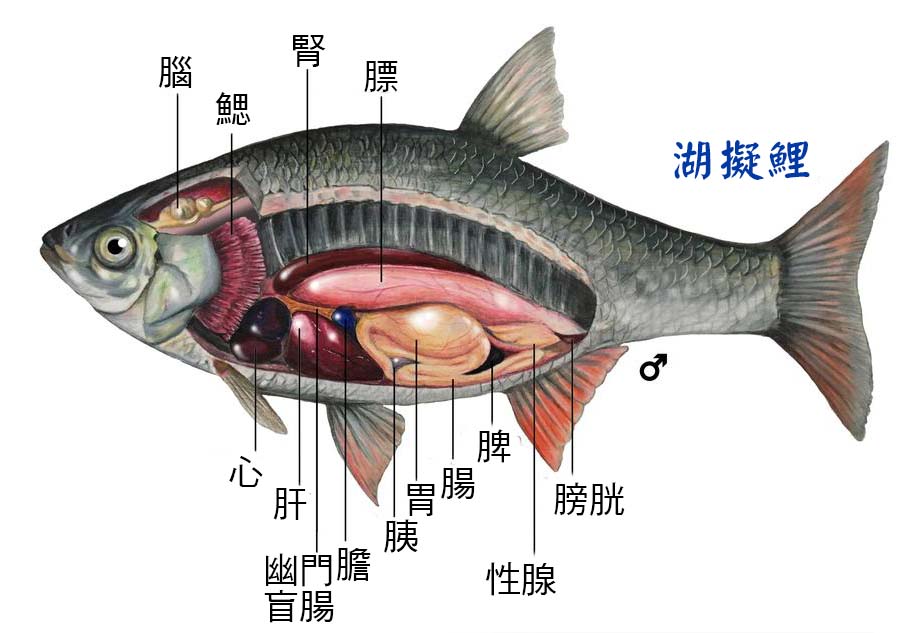
圖 2.2 魚的內部器官
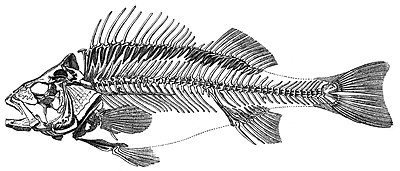
圖 2.3 硬骨魚的骨骼
2.2.2 魚類呼吸系統
魚用鰓呼吸,大多數魚類使用氣體 交換鰓上的 兩側咽, 由絲的 線狀結構組成。每 根細絲都 包含一個毛細管網絡,該網絡提供了大面積的氧氣和二氧化碳交換空間。魚通過將富含氧氣的水從口中抽出並將其 泵入鰓來 交換氣體。在某些魚類中, 毛細血管 的血液與水的流動方向相反,從而導致逆流交換。 ill 通過 咽 側面的開口將缺氧的水推出。一些魚,例如鯊魚和七 鰓鰻 ,有多個 鰻 開口。但是,硬骨魚類的兩側各有 一個鰓蓋開口 。這個開口是隱藏稱為保護 骨蓋下面 。
很多種魚可以在水中長時間生存。有些兩棲魚類,如彈塗魚、 鰻等,能離 水生活一段時間,或住在 停滯貧氧的 水體。許多這樣的魚可以通過多種機制呼吸空氣。鰻魚的皮膚可能直接吸收氧氣。肺魚類似四足動物的,必須通過 口部吞掉 新鮮空氣,並通過 鰓 傳出廢氣。許多 鯰魚讓 空氣通過腸道來呼吸。吸甲魚以利用皮膚直接吸收氧氣來呼吸 ( 類似於青蛙 ) 。也有許多魚已經進化出所謂的輔助呼吸器官,從空氣中直接提取氧氣。
居住在季節環境變化大的淺水區魚類需要直接呼吸空氣,因為該環境中水的氧氣濃度可能會季節性下降。僅僅依靠 溶解氧 的魚類會很快窒息而死,能直接呼吸空氣的魚類生存時間可以更長。在最極端的情況下,一些呼吸魚類可以在潮濕的洞穴中停水數 週 ,等待 雨季或豐水 期的到來 ( 夏季休眠 ) 。
呼吸魚類可分為專門性呼吸 和兼容性 呼吸。專門性呼吸魚類,如非洲肺魚,必須定期呼吸空氣,否則會窒息而死。 兼容性空氣 呼吸魚類,例如琵琶鼠,僅在需要時才呼吸空氣,否則將依靠 鰓 獲取氧氣。 兼容性呼吸 魚類很多,可減少抵抗水流的體力消耗和躲避掠食者及環境變化。
2.2.3 魚類血液循環系統
魚具有封閉的血液循環系統。大多數魚類的心臟位於 鰓 附近,由四個部分組成,包括 兩個腔室 和一個入口和出口。第一部分是靜脈竇,這是 一個薄壁的 囊, 從魚的 血管中收集血液,然後允許它流到第二部分,即心房,這是一個大的肌肉腔。心房用作單 向前室 ,將血液輸送到第三部分,即心室。心室是 另一個厚壁 , 肌肉腔和它 抽血,第一至第四部分,延髓動脈,大管,然後跳出心臟。動脈小動脈連接到主動脈,血液通過主動脈流到鰓進行 充氧。
2.2.4 魚類消化系統
有頜骨的 魚類 可以咬吃多種 食物,包括植物和其他生物。魚類通過口器攝取食物,並在食道中分解。 在魚胃中 ,食物被進一步消化。許多魚類,食物進入幽門盲腸的手指狀小袋加工,這 些小袋會分泌 消化 酶 並吸收營養。當食物 通過消化道時,肝臟和胰腺等器官會添加 酶 和各種化學物質。腸道完成了消化和營養吸收的過程。
2.2.5 魚類排泄系統
與許多水生動物一樣, 大多數魚將其 含氮廢物作為氨釋放。一些廢物通過 鰓 擴散。血液廢物由腎臟過濾後排放。海水魚類由於滲透作用往往會失去水分。他們的腎臟將水返回魚體本身。相反的情況發生在淡水魚中:它們傾向於滲透吸收水。他們的腎臟會 排泄稀尿 。有些魚類的腎臟經過特殊改裝,功能各不相同,可以在淡水與海水之間遷徙,特別是河口 廣鹽性 魚類。
2.2.6 魚類神經系統
與其他脊椎動物相比,魚類的大腦比例很小,通常是類似體型鳥類或哺乳動物的大腦質量十五分之一。但是,有些魚的大腦相對較大,最顯著的是鯊魚,相對於體重,它們的大腦與鳥類和有袋動物一樣大。
魚腦分為 幾個區域。 嗅球在 前面,是一對結構,通過兩個嗅覺神經接收和處理來自鼻孔的信號。 嗅葉非常 大,主要是通過 嗅味捕 獵的魚,例如鯊魚和 鯰 魚。嗅覺裂片後面是兩個 裂端腦 ,結構等同於大腦中高等脊椎動物。在魚類中, 端腦主要 與嗅覺有關。這些結構共同構成了前腦。
將前腦連接到中腦的是中腦。間腦執行與激素和 體內穩態 有關的功能。所述的松果體謊言只是間腦的上方。這種結構可檢測光線,維持晝夜節律並控制顏色變化。
在中腦 ( 或腦 ) 包含 兩個視葉 。這些種類很大,它們會視線狩獵,例如虹鱒魚。
後腦 ( 或中腦 ) 特別參與游泳和平衡。小腦是單葉結構,通常是大腦的最大部分。
腦幹 ( 或髓腦 ) 是大腦的後部。除了控制一些肌肉和身體器官外,至少在硬骨魚中,腦幹還可以控制呼吸和滲透調節。
2.2.7 魚類感覺器官
大多數魚具有高度發達的感覺器官。幾乎所有的日光魚類的色覺都至 少與人類相同。許多魚類還具有化學感受器,這些感受器會產生異常的味覺和嗅覺。儘管它們有耳,但許多魚可能聽不到很好的聲音。大多數魚類具有形成側線系統 的敏感受體,該受體能夠檢測出輕微的電流和振動,並感應附近魚類和獵物的運動。某一些魚類,例如 鯰 魚和鯊魚,具有 Lorenzini 壺腹,即電感受器,可檢測 毫伏量 級的弱電流。其他魚類,例如南美電鰻,可產生微弱的電流,將其用於導航和社交交流中。
魚類能使用地標為自己定位,並可能使用基於多個地標或符號的 思維導圖 。魚在迷宮中的行為表明它們具有空間記憶和視覺辨別力。
2.2.8.1 魚類視力
視覺是大多數魚類的重要感覺系統。魚眼鏡頭與鳥類和哺乳動物等陸地 脊椎動物的眼鏡頭相似,但晶狀體更為球形。它們的視網膜通常具有視 桿 和 視錐 ( 用於暗視和 明視覺 ) ,並且大多數物種具有色覺。有些魚可以看到紫外線,有些魚可以看到偏振光。在 無顎魚中 ,七 鰓鰻的 眼睛發達 ,而 盲鰻只有原始的眼斑。魚的視力顯示適應其視覺環境,例如深海魚類的眼睛適合黑暗環境。
魚類和其他水生動物與陸生物種生活在不同的光照環境中。 水體 會吸收 光的 波長 , 隨著深度的增加,可見光量會迅速減少。水的光學特性還會導致不同 波長 的光被不同程度地吸收,紅色、橙色等長波長的光被吸收得很快。綠色、藍色等短波長的光也會很快被吸收, 200 米以下的水體就很難有光透射入。
2.2.8.2 魚類聽力
聽力 是大多數魚類的重要感覺系統。在水下,聽力 閾 值和定位聲源的能力會降低,在這種情況下,聲速比在空中快。水下聽力是通過 骨傳導實現的 ,聲音的定位似乎取決於通過骨傳導檢測到的振幅差異。因此,水生動物很多具有在水下有效的更加專業的聽力設備。
魚可以通過其 側線 和 耳石 感覺到聲音,它們的功能就像助聽器。 鯉魚的 聽力很發達,具有 韋伯氏器官 ,這是三個專門的椎骨過程,可將游泳膀胱中的振動傳遞到內耳。
儘管很難測試鯊魚的聽力,但它們可能具有敏銳 的聽覺 ,並且可能聽到許多公里以外的獵物。頭的兩側各有一個小開口,通過細通道 直接通入 內耳 。該 側線 示出了類似的佈置,並且經由一系列開口是開放的環境稱為側線 孔 。這兩 個 振動和聲音檢測器官的組合被稱為為聲學 - 側傾系統。
壓力檢測使用韋伯 Weber 器官 ,該系統由三個椎骨附件組成,可將 氣囊 形狀的變化傳遞到中耳。它可以用來調節魚的 浮力 。像 天氣魚 和其他泥 ache 這樣的 魚 也被認為對低壓區域有反應,但它們沒有游泳膀胱。
2.2.8.3 魚類疼痛感受能力
科學家發現有些魚類有疼痛和恐懼反應。科學家的實驗中,蟾蜍魚在電擊時會發出咕嚕聲,隨著時間的流逝,它們僅在看到電極時它們就會發出咕嚕聲。虹鱒魚表現出的行為通常與其他動物的疼痛有關。蜜蜂 毒液和乙酸注入虹鱒魚嘴唇會導致魚晃動其身體,並沿水箱的側面和底部摩擦嘴唇,研究人員得出結論,這是試圖減輕疼痛的嘗試,類似於哺乳動物。神經元以類似於人類神經元模式的模式發射。
2.2.8.4 魚類運動能力
大多數魚通過在脊椎兩側交替收縮成對的肌肉而運動。這些收縮形成 S 形曲線,其向下移動到身體。當每條曲線到達 後鰭時 ,向後的力將施加到水上,並與鰭一起使魚向前移動。魚鰭的功能就像飛機的襟翼一樣。 鰭也增加 了尾巴的表面積,提高了速度。魚的流線型身體減少了與水的摩擦。由於身體組織比水更稠密,魚必須彌補差異,否則魚會下 沉 。許多硬骨魚的內部器官稱為游泳膀胱 ( 魚膘 ) ,可通過操縱氣體來調節其浮力。
硬骨魚 靠魚鰾來調節它們的 比重 , 魚藉由 魚鰾可以不用運動就緩慢上升或下降,大部分 硬骨魚類皆有 魚鰾這個調節浮力的器官。魚鰾本來是腸的一個擴充, 有些魚如 鯉魚 的魚鰾還和它的腸相連,某些 魚如 鱸 魚 的魚鰾已經 和腸完全 分開了。假如一條魚要減輕它的比重的話,它將血液中溶解的氣體釋放到魚鰾中去充氣, 鰾 也可以做為發聲共鳴的器官。軟骨魚 ( 鯊魚等 ) 及水底生活的魚沒有魚鰾,它們假如不運動的話就會 沉 到水底。
2.2.8.5 魚類吸熱能力
儘管大多數魚是變溫動物, 具有放熱性 ,但少數吸熱性的魚類 ( ,例如比目魚,鮪魚和以及 奧帕魚 (Opah) 是一種燈籠魚,可以全身吸熱,或通過游泳肌肉產生熱量加熱身體,而逆流交換 ( 如呼吸 ) 則將熱量損失降至最低。由於能夠溫暖整個身體 ( 包括心臟 ) 的能力,它能夠主動捕食魷魚等獵物並遠距離游泳。在軟骨魚類中 ,鯊科 ( 鼠 鯊 , 鯖 魚,鮭魚和大白鯊、 長尾鯊 、比目魚、藍鰭鮪魚、 海豚鯊等 ,能使體溫保持在比周圍水溫高許多的水平,強化它們的運動能力。
吸熱雖然在代謝上昂貴,但據認為具有諸如增加肌肉力量,提高中樞神經系統加工速度和提高消化速度等優點。
2.2.8 魚類生殖系統
魚的生殖器官包括 睾 丸和卵巢。在大多數物種中,性腺是大小相似的成對器官,可以部分融合或完全融合。也可能有許多次要器官會增加生殖適應性。
魚的生殖器官位於身體 側部腸的 上方。大多數魚是體外交配, 雄魚和雌 魚同時將它們的生殖細胞排泄到水中。魚卵的數量可以相差很大, 鱘魚 每次 產子可達上百萬, 而育子之的 刺魚 每次 產子不超過一百。大多數情況下養育後代的魚 中公魚照管 後代。有些魚沒有固定的性別,它們的性別隨其伴侶而變化,甚至可以在一生中多次更改。也有的魚進行體內受精,這些魚大多數直接生小魚,而不生卵。
硬骨魚類中卵的發育 過程因組而 異,對卵發生動態的確定有助於了解成熟和受精過程。在變化核, 卵質周圍 的層,以及表徵該卵母細胞成熟過程。
排卵後卵泡是卵母細胞釋放後形成的結構。它們不具有內分泌功能,具有寬闊的不規則內腔,並在涉及卵泡細胞凋亡的過程中被快速重吸收。一種稱為卵泡閉鎖的劣化過程會重新吸收未產卵的卵黃卵母細胞。這個過程也可以在其他發育階段的卵母細胞中發生,但頻率較低。
在所有已知的魚類中,有 97 %以上是卵生的,也就是說,卵在母親體外生長。卵生魚的例子包括鮭魚、鮪魚和鰻魚。在這些物種的大多數中,受精發生在母親的體外,雌雄魚的配子排入周圍的水中。但是,一些卵生魚會進行內部受精,最值得注意的是卵生鯊魚,例如 角鯊和卵生魟 魚。海魚會產生大量卵,這些 卵 通常會釋放到開放水域中。魚卵的平均直徑為 1mm 。
新孵出的 卵生魚被稱為 幼體 體 ( larvae ) 。它們通常形態不佳,帶有一個大的卵黃囊 ( 供給初期營養 ) ,與成魚外觀差異很大。卵生魚的幼體期相對較短 ( 通常只有幾 週 ) ,幼體 體 迅速生長並改變其外觀和結構 ( 稱為變態的過程 ) 成為幼體魚。在這種過渡過程中,幼體 體 必須從卵黃 囊 轉變為以浮游動物為食,這通常取決於浮游動物密度不足,使許多幼體餓死。
在卵生魚中, 卵經內部 受精後會在母親體內生長,但直接或間接地由卵母體獲得營養,而卵黃則取決於卵黃。每 個 胚胎都在自己 的卵中發育 。卵生魚的常見例子包括孔雀魚,天使鯊和 腔棘 魚。
有些魚類是胎生的。在這樣的物種中,母親保留卵子並滋養胚胎。通常,胎生魚的結構類似於在哺乳動物中看到的胎盤,該胎盤將母親的血液供應與胚胎的血液連接起來。胎生魚類的例子包括衝浪 鱸 , 裂鰭和檸檬鯊。一些胎生魚表現出卵形,正在發育的胚胎會吃掉母親生產的其他卵。
2 .2.9 魚的聲音交流
魚中的聲音交流涉及將聲音信號從一個物種的一個個體傳輸到另一個物種。聲音的產生是魚類之間交流的一種手段,最常用於餵養,侵略或求愛行為的背景下。 發出的聲音會根據所涉及的物種和刺激而有所不同。魚可以通過移動骨骼系統的成分來產生刺耳的聲音,也可以通過操縱專門的器官 ( 例如游泳囊 ) 來產生不刺耳的聲音。
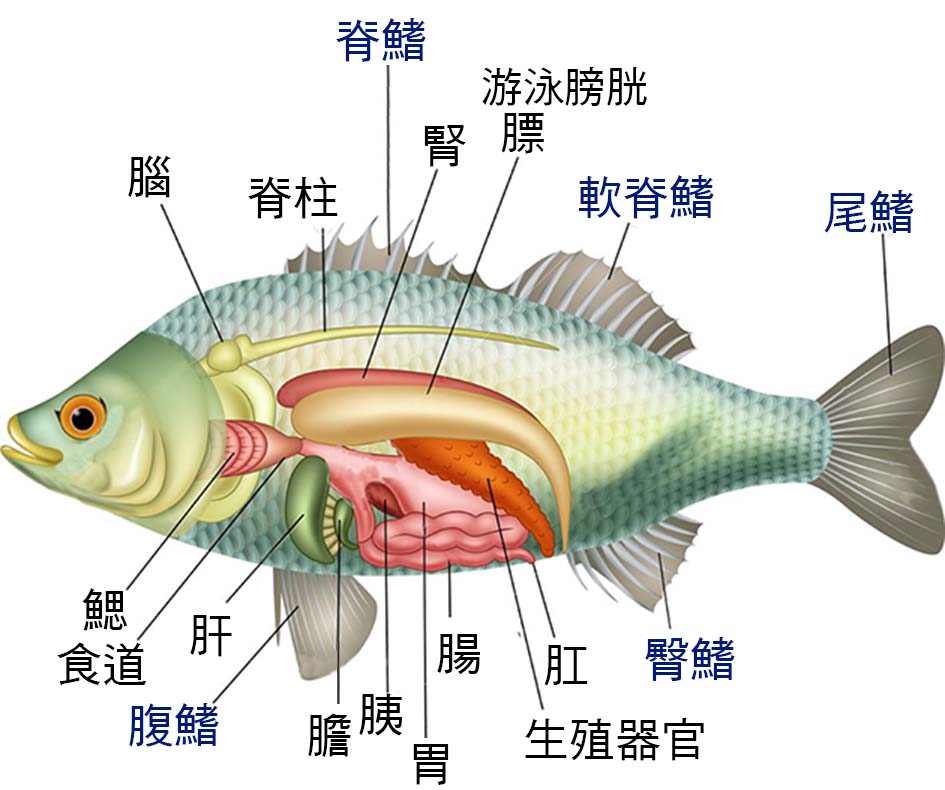
圖 2.4 魚的構造
2.3 魚的外部器官
魚類的身體結構通常取決於水的物理特性,水是魚賴以生存的媒介。水比空氣密度大得多,其溶解的氧相對較少,並且吸收的光比空氣多。魚的身體分為頭部, 軀乾和 尾部,儘管三者之間的劃分並不總是在外部可見。形成魚內部支撐結構的骨骼是由軟骨 ( 軟骨魚 ) 或硬骨 ( 硬骨魚 ) 製成的。主要的骨骼元素是椎骨柱,由關節椎骨組成重量輕但堅固。肋骨附著在脊柱上,沒有四肢或四肢帶。魚的主要外部特徵是 鰭 ,由稱為射線的骨刺 或軟刺組成 ,除尾鰭外,與脊柱沒有直接聯繫。它們由構成軀幹主要部分的肌肉支撐。。眼睛適合在水下看,只有局部視野。有一個內耳但沒有外耳或中耳。低頻振動是由沿著魚兩邊長度的感官器官的側線系統檢測到的,該系統對附近的運動和水壓的變化做出響應。
鯊魚和 魟 魚是較原始魚類,具有許多與古代魚類相似的原始解剖特徵,包括由軟骨組成的骨骼。他們的身體傾向於背 腹展平 ,他們通常在頭的下面有五對 鰓縫和 一個大嘴巴。在真皮層上覆蓋有獨立的真皮盾 鱗 。它們有一個泄殖腔,尿道和生殖器通道 可通入其中 ,而膀胱則不行。軟骨魚類產生少量的大型卵。一些種類是卵生的,具有年輕的內部開發,但其他人是卵生和幼體在外部發展卵鞘。
硬骨魚的譜系顯示出更多的衍生解剖特徵,它們具有骨架,一般都是橫向扁平,具有 5 對由保護鰓 厴 ,和一個在口或口 鼻部的尖端 附近。真皮覆蓋著鱗片。硬骨魚有一個 游泳膘囊 ,可以幫助它們在水體中保持恆定的深度,但不會使泄殖腔保持恆定。他們大多產卵大量蛋黃的小卵, 然後撒入 水體中。
2.3.1 椎骨
魚是脊椎動物。所有脊椎動物均按照基本的碳酸鹽體計劃建造:一根堅硬的 桿 貫穿動物的整個長度 ( 椎骨或脊索 ) ,上方有一根神經組織 ( 脊髓 ) 的空心管,下方是胃腸道。在所有脊椎動物中,嘴位於動物的前端或在動物的前端下方,而肛門則在機體末端之前向外張開。肛門以外的身體其餘部分形成有椎骨和脊髓的尾巴,但沒有腸。
脊椎動物的定義特徵是椎骨柱,其中在所有脊索中發現的脊索已被由活動關節 ( 椎間盤 ,胚胎起源 ) 分隔的一系列較硬的椎骨所代替並從脊索進化而來 ) 。
脊柱由中樞的頂部和底部突出的脊椎 弓 以及從中樞 或弓突出 的各種過程組成。從中樞的頂部延伸 的弓被稱為 神經弓。
在包括所有硬骨魚在內的大多數 輻 鰭魚 中,這兩 個 結構與一塊堅固的骨頭融合在一起並嵌入其中,這些骨頭表面上類似於哺乳動物的椎體。在鯊魚等軟骨魚類中,椎骨由兩個軟骨管組成。 上管由 椎弓形成,還包括填充在椎骨之間的間隙中的附加的軟骨結構,將脊髓包圍在基本上連續 的鞘中 。 下管圍繞 著脊索並具有複雜的結構,通常包括多層鈣化層。
2.3.2 魚頭
魚類頭部或顱骨包括顱骨頂 ( 一組覆蓋大腦,眼睛和鼻孔的骨頭 ) ,鼻 ( 從眼睛到 上頜骨的 最前端 ) 、 鰓蓋 ( 鯊魚 沒有鰓蓋 ) 和臉頰,從眼睛延伸到前鰓、 鰓蓋可能 沒有刺。
魚的頭骨是由一系列僅鬆散連接的骨頭組成的。 無顎魚和 鯊魚僅具有軟骨內顱骨,而軟骨魚的上顎和下顎是 不 附著在頭骨上的獨立元素。硬骨魚有額外的真皮骨頭。
頭部可能有幾種肉質結構,稱為槓鈴,可能很長,類似於鬍鬚。許多魚類的頭部也有各種突起或刺。幾乎所有魚類的鼻孔或鼻孔都沒有連接到口腔,而是形狀和深度不同的 凹 坑。
2.3.3 魚下顎 jaw
脊椎動物的下顎可能最初是在志留紀時期演化的,並出 現在 盾皮魚綱 中,在泥盆紀中進一步多樣化。人們認為下頜起源於支撐魚鰓 的咽弓 。 這些弓 的最前面的兩個被認為是 頜骨本身和舌骨弓 , 將頜骨支撐在腦箱上 ,提高了機械效率。
許多魚類,下頜獲得的原始選擇優勢與進食無關,而與提高呼吸效率有關。顎 式幫浦將 水跨過魚類或空氣抽入兩棲動物的肺部。當然魚下頜也能幫助進食,並在脊椎動物中成為非常重要的功能。
2.3.4 魚眼睛
魚眼類似於鳥類和哺乳動物等陸地脊椎動物,但具有更球形的透鏡,如同照相機中魚眼鏡頭的效果,可以觀察很寬的超廣角。它們的視網膜通常同時具有桿狀細胞和 視錐 細胞 ( 用於暗視和 明視覺 ) ,並且大多數魚類具有色覺。有些魚可以看到紫外線,有些魚可以看到偏振光。在 無顎魚中 ,七 鰓鰻的 眼睛很發達,而肺魚只有原始的眼點。它們被推到很深的黑暗水域,在那裡它們不易被肉眼看到的掠食者所吸引,並且在這裡最好有一個 凸 眼點,它比平或 凹 眼 點集光 更多。與人類不同,魚通常通過將晶狀體移 近或移離 視網膜來調整焦點。
2.3.5 魚鰓 gill
位於鰓蓋下 的魚鰓是呼吸器官,用於從水中提取氧氣和排泄二氧化碳。它們通常不可見,但在某些物種中可見,用於允許 魚從空氣提取氧。耙是指 在 鰓弓上 的手指狀突起,在過濾器進料器中起到保持過濾獵物的作用。它們可能是硬骨魚的或軟骨魚。
2.3.6 魚表皮
魚的表皮完全由活細胞組成,在表層細胞中只有極少量的角蛋白。它通常是可滲透的。硬骨魚的真皮通常只含有很少的四足動物結締組織。相反,在大多數魚類中,它已被堅固的保護性骨鱗片所代替。除了一些特別大的真皮骨 ( 構成頭骨的一部分 ) 外,這些鱗片也消失在四足動物中,儘管許多爬行動物的確有不同種類的鱗片,而穿山甲也是如此。軟骨魚類有許多像牙齒一樣的細齒嵌入自己的皮膚來代替真正的尺度。
汗腺和皮脂腺都是哺乳動物獨有的,但是魚類中也發現了其他類型的皮膚腺。魚通常有眾多的個人粘液分泌型的皮膚細胞,目的為絕緣和保護,但也可能有毒液的腺體,或細胞漿液。黑色素使許多物種的皮膚著色,但是在魚類中,表皮通常相對無色。相反,皮膚的顏色很大程度上是由於真皮 中的色譜產生 的,除了黑色素,還可能含有鳥嘌呤或類胡蘿蔔素色素。許多物種,例如比目魚,通過調整其 色譜圖 的相對大小來改變其皮膚的顏色。
2.3.7 魚鱗片 scales
許多魚的外部被鱗片覆蓋,這是魚外皮系統的一部分。鱗片起源於中胚層 ( 皮膚 ) ,其結構可能與牙齒相似。一些魚類被覆蓋鱗甲代替。也有些魚類的皮膚上沒有外殼。大多數的魚類被粘液 ( 粘液 ) 保護層覆蓋。鱗片的另一種較不常見的類型是骨板。一種經過修飾的增厚的鱗片,通常是龍骨狀或多刺的。
2.3.8 魚側線
魚身體上的側線是用於檢測周圍水中的運動和振動的感覺器官。例如,魚類可以利用其側線系統追蹤 逃逸獵物產生的渦流。在大多數物種中,它由沿著魚兩邊延伸的一排受體組成。
2.3.9 螢 斑 Photophores
許多魚類外觀有部分閃耀的螢光,螢光體是發光的器官,在某些魚類上以發光點的形式出現。 光線可以由獵物消化過程中的化合物產生,也可以由稱為光細胞的有機體中專門的線粒體細胞產生,也可以由共生細菌產生。 螢光粉用於吸引食物或使捕食者迷惑。
2.3.10 魚鰭 fins
魚鰭是魚的最鮮明特徵。它們或是由骨刺構成,或是從身體突出的 輻鰭線 構成,覆蓋魚的皮膚,然後將它們連接在一起,就像大多數硬骨魚所看到的那樣是網狀,或者類似於鯊魚中的鰭狀 肢 。除了尾鰭之外,幾乎所有的魚鰭都與脊椎柱沒有直接連接,僅由肌肉支撐。它們的主要功能是幫助魚游泳。 鰭也可 用於滑行或爬行,如飛魚和能爬行的天使魚。 位於魚上不同 位置 的鰭有不同 的用途,例如向前移動,轉向和保持直立姿勢。對於每 個鰭 ,都有許多魚類在進化過程中失去了這種特定的 鰭 。
2.3.11 魚刺和 輻鰭線 Spines and rays
在硬骨魚中,大多數 鰭 可能有刺或 輻鰭線 。 輻 鰭通常是柔軟的、有彈性的、分段的。 輻鰭線 的這種分割是將 它們與刺區別 開的主要區別。
魚刺有多種用途。在 鯰 魚中,它們被用作防禦的一種形式。許多 鯰 魚有能力將其脊椎向外鎖定。甚至使用外骨刺將自己固定在岩石縫隙中,以防止它們被拉出。
鱗翅目魚類是在硬骨魚中發現 的雙側成對 的骨鰭 鰭 。它們在 輻鰭線 周圍發展,是真皮外骨骼的一部分。
2.4 魚鰭的類型
• 背鰭:背鰭位於魚背上,可防止魚滾動,並有助於突然的轉彎和停止。大多數魚有一個背鰭,但有些魚有兩個或三個。
• 尾鰭 / 尾鰭:也稱為尾鰭,尾鰭附著 在尾柄的 末端,用於推進。 尾柄是 魚身體的狹窄部分。脛骨關節是尾鰭和最後一個椎骨之間的關節。尾 輻 鰭通常是扇形的。
• 肛門 鰭 :位於肛門後腹面,用於在游泳時穩定魚身體的 鰭 。
• 胸鰭:兩側成對,通常在蓋後面。胸鰭與四足動物的前肢同源,並有助於在一些魚類中行走,例如一些澳洲天使魚。在某些魚類中高度發達的胸鰭的特殊功能是動態提升力的產生,該動態提升力有助於諸如鯊魚之類的某些魚類保持深度,並能使飛魚滑翔。胸鰭的某些 輻 鰭可以適應手指狀的投射,例如飛魚。
• 骨盆鰭 / 腹鰭:骨鰭位於四肢下方,在腹鰭 的腹側各 成對。它們幫助魚在水中上下移動,急轉彎並迅速停止。在 鰕虎 中,骨盆鰭通常融合在單個吸盤中,該吸盤可用於附著到物體上。
• 脂肪 鰭 :柔軟的肉質 鰭 ,位於背鰭後面和尾鰭的正前方。它在許多魚類科目中都不存在,但在 鮭 科、 擬脂鯉和鯰 魚中發現。它的功能仍然 是個謎 ,相關研究表明, 鰭片對於 檢測和響應諸如觸摸,聲音和壓力變化的刺激可能至關重要。
• 尾龍骨:通常在尾鰭前面 的尾柄上 的側脊,某些類型的快速游動的魚類上,它像魚龍骨一樣為尾鰭提供穩定性和支撐。可以有一對龍骨,每側一個,或者上下兩對。
• 鰭片 : 小鰭片通常 位於背鰭和尾鰭 之間, 但也可能位於 肛門鰭和尾鰭 之間。在雙足動物中,背鰭上只有 鰭 ,而背鰭沒有。在一些鮪魚或秋刀魚等魚類中,它們是無射線的,不可縮回的,並且位於最後的背鰭或 肛門鰭與尾鰭 之間。
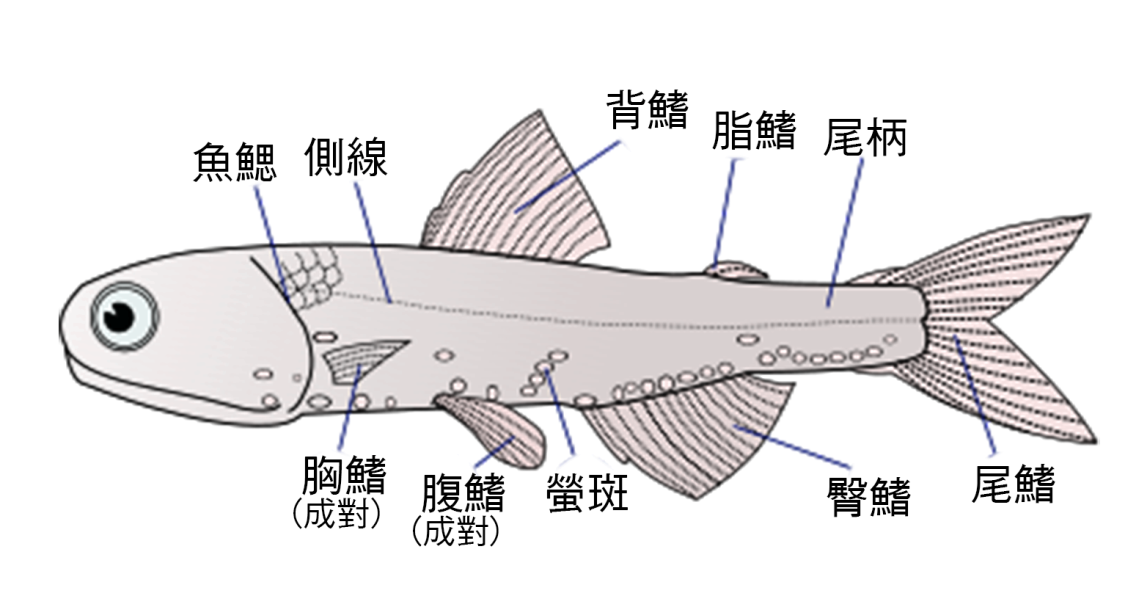
圖 2.5 魚鰭名稱與位置
2.5 魚的內部器官
2.5.1 魚腸
與其他脊椎動物一樣,魚類的腸道由兩部分組成,即小腸和大腸。在大多數較高等的脊椎動物中,小腸進一步分為十二指腸和其他部分。在魚類中,小腸的劃分不 清楚, 可以使用術語 前腸或近端腸 代替十二指腸。 在硬骨魚中,腸相對較短,通常約為魚體長度的一半。它通常有一些幽門 盲腸沿其長度 的小袋狀結構,有助於增加用於消化食物的器官的總表面積。硬骨魚中沒有 迴盲閥 ,小腸和直腸之間的邊界僅以消化上皮的末端為標誌。在非成鰭魚類中,如鯊魚和肺魚,也沒有小腸。取而代之的是,腸道的消化部分形成螺旋狀的腸子, 連接胃到直腸 。在這種類型 的腸中 ,腸本身是相對筆直的, 但是沿內表面 呈螺旋狀延伸較長的褶皺,有時彎曲數十圈。極大地增加了腸的表面積和有效長度。在硬骨魚和非哺乳動物的四足動物中,螺旋腸的 內襯與小腸 的內襯相似。
2.5.2 魚幽門與盲腸
魚的幽門盲腸在大腸開始處是一個袋,通常是腹膜的。它從 迴 腸接收糞便,並連接到大腸的升結腸。許多魚類的腸內有許多小東西,也叫幽門盲腸。它們的目的是增加消化上皮的總表面積,從而 優化糖 ,氨基酸和二 肽 以及其他營養物質的吸收。
2.5.3 魚胃
大多數硬骨魚類與其他脊椎動物一樣,食道和十二指腸開口到胃的相對位置保持相對恆定。結果,胃在彎曲回幽門括約肌之前總是向左彎曲。然而,七 鰓鰻、盲鰻 、肺魚和少數硬骨魚根本沒有胃,食管直接通向腸道。這些魚的飲食需要很少的食物儲藏,或者不需要預先用胃液消化。
2.5.4 魚腎臟
魚的腎臟通常是狹窄的細長器官,佔據了軀幹的很大一部分。它們是 類似中腎的 高等脊椎動物。腎臟內有 一群腎 單位,由收集通常排 入中腎導管 的導管服務。但是,情況並不總是那麼簡單。在軟骨魚中,還有一條較短的導管,該導管排出腎臟的後部分,並在膀胱或泄殖腔處 與中腎導管 相連。
2.5.5 魚脾臟
脾臟在幾乎所有脊椎動物中都能發現。它是非重要器官,其結構類似於大淋巴結。它主要充當血液過濾器,在紅細胞和免疫系統方面起著重要作用。在軟骨和硬骨魚中,它主要 由紅漿組成 ,並且通常是稍微拉長的器官,因為它實際上位於腸的漿膜內壁。僅有脾臟的無脊椎動物是七 鰓鰻和盲鰻 。即使在這些動物中,在腸壁內也有 一個造 血組織的擴散層,其結構與 紅漿相似 。推測與高級脊椎動物的脾臟同源。
2.5.6 魚肝臟
肝臟是所有魚類中存在的 重要重要 器官。它具有廣泛的功能,包括排毒,蛋白質合成和消化所需的生化試劑的生產。它很容易受到有機和無機化合物的污染,因為它們會隨著時間的推移而積累,並可能危及生命。由於肝臟具有排毒和儲存有害成分的能力,因此常被用作環境生物標誌物。
2.5.7 魚心臟
魚具有通常所 說的兩腔心 ,由一個中庭接收血液和一個 心室泵血 , 與兩室兩棲動物 ( 兩個心房,一個心室 ) 組成,兩棲動物最爬行。心臟和哺乳動物和鳥類心臟的 四個腔室 ( 兩個心房,兩個心室 ) 。但是,魚心有進入室和出口室,這些室可能 稱為腔室 ,因此根據 算作腔室 的不同,有時也將其描述為三 腔室或 四 腔室 。心房和心室有時被認為是 “ 真腔室 ” ,而其他的則被認為是 “ 附屬腔室 ” 。
四個隔間按順序排列:
1. 靜脈竇:帶有一些心肌的 薄壁囊或水庫 ,可通過進入的肝靜脈和心臟靜脈收集脫氧的血液。
2. 心房: 壁較厚 的肌肉腔,將血液輸送到心室。
3. 心室: 厚壁的 肌肉腔,將血液泵送到第四部分,即流出道。腦室的形狀變化很大,通常在帶有細長體的 魚中呈管狀 ,在其他 魚中呈三角形 ,具有三角形底,或者在某些海魚中有時 呈囊狀 。
4. 流出道 (OFT) :轉到腹主動脈和由管狀的動脈圓錐,延髓動脈,或兩者。動脈圓錐通常在較原始的魚類中發現,它收縮以幫助血液流向主動脈,而腓骨前動脈則沒有。
由瓣狀 結締組織組成的 骨瓣可 防止血液倒流通過隔室。 竇 靜脈和心房之間 的耳腔瓣膜 稱為 竇房閥 ,在心室收縮過程中會關閉。在心房和心室之間的是一個稱為房室瓣的小動脈瓣膜,而在 小球房和 心室之間的是一個稱為 小腦室瓣的 小動脈瓣。動脈圓錐具有可變數量的 半月瓣 。
在成年魚類中,四個隔間不是成一直線排列,而是形成 S 形,後兩 個 隔間位於前兩 個 隔間的上方。在軟骨魚和魚鰭魚中發現了這種相對簡單的模式。在 硬骨中 ,動脈圓錐非常小,可以更準確地描述為主動脈的一部分,而不是心臟的一部分。羊膜中沒有動脈圓錐,大概是在進化過程中被吸收到了腦室中。同樣,在一些爬行動物和鳥類中,竇靜脈呈殘留結構,否則會被吸收到右心房中,不再可分辦。
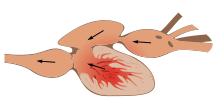
圖 2.6 魚類心臟模型
2.5.8 游泳膀胱 ( 魚膘 )
游泳膀胱或氣囊是一種內部器官,有助於魚類控制浮力,從而保持在當前水深,上升或下降而不必在游泳中浪費能量。僅在硬骨魚中發現了膀胱。鮪魚和 鯖 魚等快速游泳的魚類通常不存在這種 膀胱魚膘 。
2.5.9 韋伯裝置
許多魚類具有一種稱為韋伯裝置 (Weberian) 的結構,這種結構可以使它們聽到更好的聲音。這種能力由一組稱為韋 伯小骨 的骨頭組成,這是一串小骨頭,將聽覺系統連接到魚類的游泳膀胱。這允許振動傳遞到內耳。
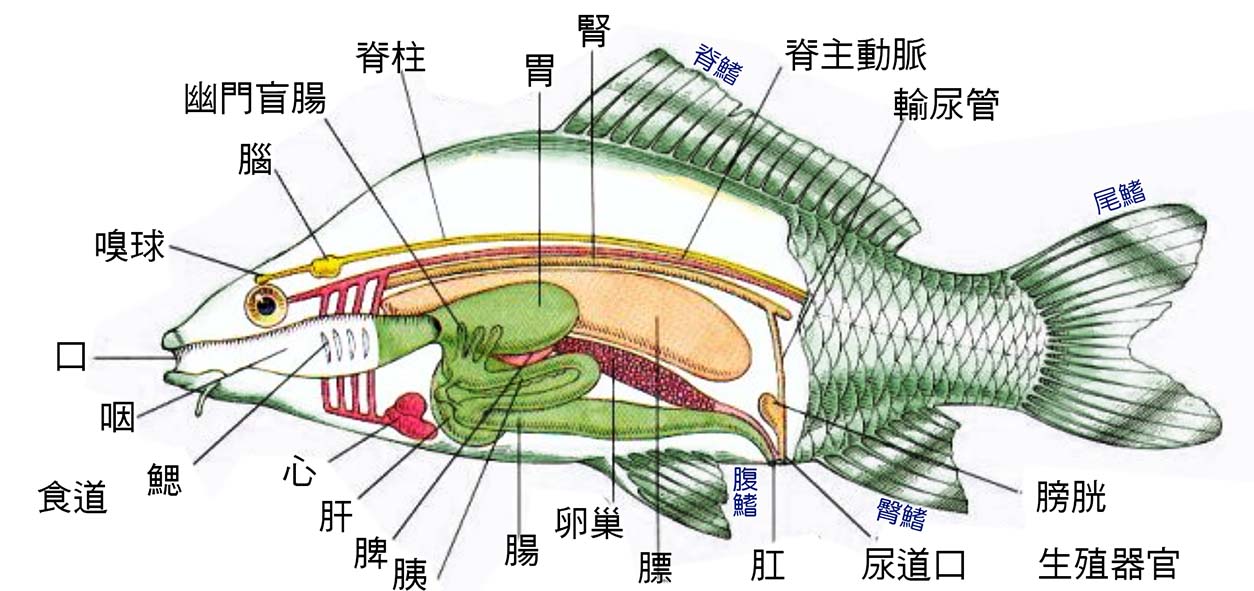
2.6 魚的疾病與免疫
免疫 器官因魚的 類型而異。在 無顎魚 ( 七 鰓鰻和盲鰻 ) 中,沒有真正的淋巴器官。這些魚依靠其他器官內的淋巴組織區域來產生免疫細胞。
軟骨魚類 ( 鯊魚和 魟 魚 ) 具有更先進的免疫系統。他們有軟骨魚特有的三個專業器官 ; 圍繞性腺的生殖器官 ( 類似於哺乳動物骨骼的淋巴組織 ) ,食道壁內 的間質細胞 (Leydig) 以及腸中的螺旋瓣。這些器官容納典型的免疫細胞 ( 粒細胞,淋巴 細胞和漿細胞 ) 。它們還具有可識別的 胸腺和發育 良好的脾臟 ( 最重要的免疫器官 ) ,其中發育並儲存著各種淋巴細胞,漿細胞和 巨噬 細胞。
軟骨魚類在與腦膜 ( 中樞神經系統周圍的膜 ) 相關 的團塊內 具有 生產粒 細胞的主要場所。他們的心臟經常覆蓋著含有淋巴細胞,網狀細胞和少量巨噬細胞的組織。軟骨骨骼腎臟是重要的造血器官。它是紅細胞,粒細胞,淋巴細胞和 巨噬 細胞發育的地方。
硬骨魚類的免疫系統,包括腎 ( 尤其是 前腎 ) ,裡面有很多不同的免疫細胞。 此外, 硬骨魚在黏膜組織內 ( 例如皮膚,腮, 腸和性腺 中 ) 具有胸腺,脾臟和分散的免疫區域。與哺乳動物的免疫系統很像, 硬骨紅 細胞,嗜中性粒 細胞和粒細胞 被認為存在於脾臟中,而淋巴細胞是在 胸腺中發現 的主要細胞類型。
2.6.1 魚類疾病和寄生蟲
像其他動物一樣,魚也患有疾病和寄生蟲。為了預防疾病,他們有多 種防禦措施。非特異性防禦包括皮膚和鱗屑,以及表皮分泌的粘液層,該粘液層可以捕獲並抑制微生物的生長。如果病原體破壞了這些防禦能力,魚會產生發炎症反 應,從而增加血液流向感染區域並輸送白細胞,從而破壞病原體。特定的防禦措施對魚類身體識別的特定病原體有反應,即免疫反應。近年來,疫苗已廣泛使用於養 殖漁業、觀賞魚。
一些物種使用清潔的魚類去除外部寄生蟲。有名的最好的是發現的印度和太平洋的海洋珊瑚礁藍霹靂清潔工 瀨魚屬 ( Labroides ) 。這些小魚擁有所謂的 “ 清潔站 ” ,其他魚聚集在那裡並進行特定的運動以吸引清潔工的注意。在許多魚群中都觀察到了清潔行為,包括一個有趣的案例,即同一屬的花斑 腹麗魚 ,也是專門幫魚類清除寄生蟲的魚醫生。