
魚類生態學
1. 水環境與魚分類
2. 魚類生理學
3. 環境條件對魚種分佈的影響
4. 生物因數與魚類群落結構
5. 移棲地 與魚群行動
5.1 概述
5.2 游泳能力和能源成本
5.3 模式和 棲地 依戀與社交互動
5.4.1 地域範圍和領域感
5.4.2 群聚 Shoal ing
5.4 遷徙
5.5.1 遷徙的定義
5.5 河流中的魚類活動
5.6.1 本地範圍和領域
5.6.2 河流內遷移
5.6 湖泊的社會結構和運動
5.7.1 社會結構
5.7.2 湖泊內遷移
5.7 珊瑚礁和 岩岸 的社會結構和運動
5.8 海洋魚類的社會結構與運動
5.8.1 遠海魚類
5.8.2 遠海魚類的遷徙
5.8.3 遠海魚類的垂直遷移
5.8.4 底棲魚類的社會結構和運動
5.9 迴游 school ing
5.9.1 迴游 分佈
5.9.2 溯河魚類的社會結構
5.10 返鄉 Homing
5.10.1 返鄉機制
5.11 撈捕的影響
6. 攝食與生長
7. 生活史和種群動態
8. 魚類應用生態學
第五章 魚類的遷徙、屬地性和淺水化
5.1 概述
水的運動範圍從山間溪流的急流、海浪對岩石海岸的撞擊、潮汐在平緩擱淺的海灘上的漲落,到緩慢移動的洋流。除非魚表現出積極的行為機制來阻止它,否則它們會隨著水流一起移動。卵、幼體和 幼魚 要麼缺乏主動運動的機制,要麼這些機制發育得太差,無法防止被動移位。在生命史的早期階段,魚類的活動很大程度上取決於卵在何時何地下蛋。隨著運動能力的成熟,魚變得能夠抵抗水的運動,並且能夠對它們將生活的棲息地做出積極的選擇 ( 第 2 章和第 3 章 ) 。
這些選擇可以從兩個維度來描述: ( i ) 個體魚類對給定地點的依附性;和 (ii) 與同一物種的其他魚類 ( 同種相互作用 ) 和與其他物種的社會相互作用的性質 ( 異種相互作用 ) ( 表 5.1) 。在許多物種中,個體魚在其一生中會改變它們的行為,因此也會改變它們在這兩 個 維度上的分類。在更詳細地探討這一分類之前,考慮魚類的遊動能力與其大小的關係以及在不同速度下游泳的能量消耗是有用的。
表5.1 基於兩個維度的運動方式分類:場所依戀和社會互動性

5.2 游泳能力和能源成本
前面章節描述了魚類形態與棲息地的關係。體型是影響游泳能力的另一個主要因素。大小影響魚的操作和可以達到的絕對速度。
硬骨魚幼體 在其水動力 環境中 經歷了粘性 阻力。它的作用是,運動一旦停止,幼魚就不能向前滑行而不主動游泳 (Webb 和 Weihs , 1986) 。對於較大的魚,粘性阻力變得不那麼重要,而壓力阻力占主導地位 ( 表 1.2) 。在這種水動力狀態下,大多數幼魚和所有成年魚基本上都佔據著這種狀態,這種動物在一段主動推進期結束時可以在水面上滑行。這種突 入式和海岸 式游泳的特點是身體和 鰭的 一個或多個運動週期,加速魚的運動 ( 爆裂 ) ,然後是一個沒有推進運動的滑行階段。這種魚獲得了一個優勢,因為在滑行階段,它可以保持身體在一條直線上,這將阻力最小化。在推進階段,身體必須彎曲,這會增加阻力。
當討論游泳速度、體型和游泳的能量消耗時,定義三個速度範圍是有用的:持續游泳、長時間游泳和爆發游泳 (Hoar 和 Randall, 1978) 。在持續游泳中,游泳肌肉所需的能量由有氧呼吸提供。魚不會 積累氧債 ,只會慢慢疲勞。游泳速度通常被限制在每秒 6 到 7 個身體長度以下,在長時間的游泳比賽中通常保持較低的速度。在爆發式游泳中,可以達到更高的速度,有時高達每秒 20 個身長。然而,爆發式游泳的能量是由無氧呼吸產生的,疲勞非常迅速。長時間游泳是介於爆發式游泳和持續游泳之間的一種中間類型。能量由有氧和無氧代謝提供。長時間游泳可能持續 15 到 3 小時,但長時間的游泳最終會導致疲勞。捕食者在短距離的水域中追逐獵物 ( 或魚網 ) ,以快速遊動,以顯示其快速遊動的能力。維持游泳的能力對於大面積覓食、在緩慢但穩定的水流面前保持姿勢或長距離遷徙非常重要。為了比較不同大小或種類的魚,通常使用臨界游泳速度。 當魚在 深水潭中逐漸增加游泳速度時,達到疲勞前的最大速度。
最大速度隨著魚的大小而增加。這種增長通常可以用以下關係來描述:
log V = a + b log L
式中 , V= 遊動速度 (mm/sec) 和 L 為魚體長度 (mm)(Beamish, 1978) 。例如,一條長 100mm 的大 馬哈魚的 臨界速度約為 600mm/sec( 每秒 6 條體 長 ) ,但在 500mm 時,其臨界速度約為 1400mm/sec 。當游泳速度是以身體長度而不是每秒 mm 來衡量時,較小的魚往往比較大的魚表現更好,儘管它們的絕對速度較低。
游泳是一種耗資巨大的運動方式 (Schmidt-Nielson, 1984) 。衡量這一成本的一個指標是將一克魚移動一公里 (/ Jg /km) 所消耗的能量。這種成本隨著魚的尺寸的增加而降低,但是對於目前為止研究過的那些物種來說,同樣大小的魚的成本是相似的。對於體重為 10 克的紅 鮭 ,每公里的能量消耗約為 46 焦耳 (4.6/ Jg /km) ,但對於體重為 1000 g 的紅 鮭 ,其成本約為 1430 焦耳 (1.4/ Jg /km)(Beamish, 1978 ; Schmidt-Nielson, 1984) 。較大的魚類可能比較小的魚類更容易進行長途遷徙,因為前者的旅行效率比後者高 ( Roff 1988) 。這些數值是指持續游泳。突然游泳要貴得多。對於幼年銀 鮭 來說,爆發式游泳在能量方面比以最大可持續速度游泳的成本高出近 40 倍 (Puckett 和 Dill 1984) 。能否逃脫捕食者的突襲是一個生死攸關的問題;移動的速度比移動的效率更重要。
5.3 棲息地依戀和社交互動模式
5.3.1 領域感和歸屬範圍
釣魚者有時會 “ 編造 ” 在已知地點潛伏的特殊魚類的故事。這些意見是指觀察出現魚 蹤 概率的概念。魚有游泳能力,但是魚群也會有領域感,個體的全部或部分生命都生活在固定棲息地。 Gerking (1959) 。
在某些物種中,個體魚表現出位置的依附性,但不同個體所佔據的區 域可能重疊,並且沒有個體將另一個體排除在特定區域之外的行為機制。每群魚都佔據一個家的範圍。居住在一個家庭範圍內可能會讓魚瞭解食物和住所的位置,在 某些情況下,還有適合繁殖的地點。如果一個地區的魚群可以將其他魚排除在該地區之外,那麼該居民就是在宣示領土主權。地形行為使居住者能夠自行扣押該地區 的資源。為了保持對其他魚類的排斥而必須支付的任何費用必須抵消這種隔離的好處。該區域可能只防禦同種異體,或在某些例子中同時對抗異種魚類 。 被隔離的資源可能是食物、庇護所、產卵地或這些的組合。
5.3.2 群聚
魚一生中的一部分時間,可能與群聚點中的其他魚類一起度過。魚群的形成取決於魚在社會上的相互反應,而不僅僅是魚聚集在對它們有吸引力的地點,而不是因為與其他魚類的存在無關的地方。術語 “ 群聚 shoaling ” 和 “ 魚群巡 游 schooling ” 有時可以互換使用,但對於魚群中的魚類在魚群內的空間位置上表現出高度的協調性以及良好的同步動作,保持群 游 是有用的 (Pitcher, 1986) 。
群聚或群 游中的魚被捕食 者攻擊的風險可能降低 (Pitcher, 1986) 。它還可以提高個體找到食物的機會,食物分散在整個棲息地中。在魚群中生存的一個可能的代價是,當發現食物時,對食物的競爭會加劇,因為魚群的成員身份確保了個體被其他個體包圍。
在某些物種中,群聚可能覆蓋廣闊的區域,而在其他物種中,群聚可能佔據一個家的範圍。單 個 群聚對空間的利用是一個迫切需要更多資訊的主題。
5.4 遷移
5.4.1 遷徙的定義
許多群聚魚種也會遷徙。遷移的一個有用的定義是那些導致兩個更為獨立的棲息地之間交替的、有規律的週期性發生並涉及大量種群的遷移 (Northcote, 1978 年 ) 。這種魚在一生中在棲息地之間來來往往,要麼一次,要麼週期性地 ( Roff , 1988) 。 為了進行洄游,魚類必須失去它們可能擁有的、在洄游階段結束時可能再次承擔的任何地點附著物資源。根據種群豐度來衡量,許多遷徙物種都非常成功,這表明, 就繁殖成功而言,遷徙行為的好處大於不可避免的代價。這些成本將包括移動的能源成本和由於魚類通過不熟悉的地點而增加捕食的可能性。這些好處是累積的,因 為這些運動 將魚帶到 比它們離開的棲息地更適合它們的棲息地。
通常,遷徙會在適合覓食的棲息地和適合繁殖的棲息地之間捕食 ( 圖 5.1) 。它們還可能包括從棲息地進出的活動,這些棲息地形成了對環境不利的庇護所,這些環境要麼是由於不利的非生物條件,要麼是因為捕食者或病原體的存在使它們變得危險。描述魚類遷徙的術語現在已經發展出來了 (Harden Jones, 1968 ; McDowall, 1988) 。
河豚類的遷徙完全局限於淡水。它們可能涉及河流系統內的遷移或河流和湖泊之間的遷移。遠海物種完全在海洋系統內遷移。許多最重要的漁業撈捕物種都是遠海物種。溯河物種在海洋和淡水系統之間遷移 ( 圖 5.2) 。分為三類。溯河產卵的物種遷移到淡水中產卵,但他們的一部分生命都在海裡覓食。 降海產卵 物種遷徙到海裡產卵,但在其生命週期的一部分時間裡都在淡水中覓食與成長。兩棲類物種在海洋和淡水之間遷移,但遷移與繁殖沒有直接聯繫。溯河物種的特點是它們具有在具有 不同岩杜滲透性 和離子性質的水域之間移動的生理能力。
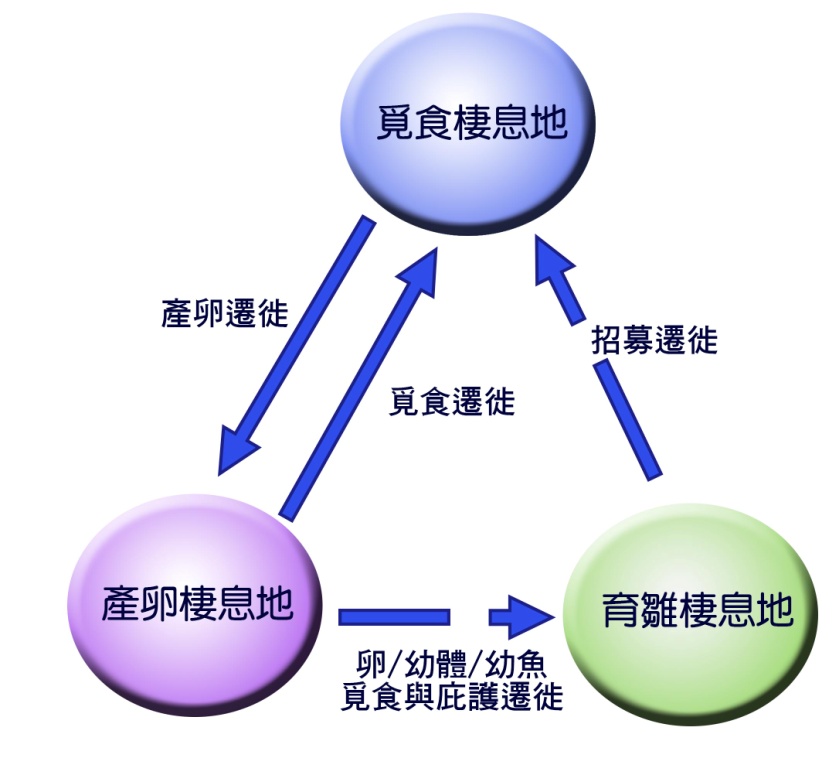
圖 5.1 餵養,產卵和 育雛 棲息地之間遷移的基本模式。
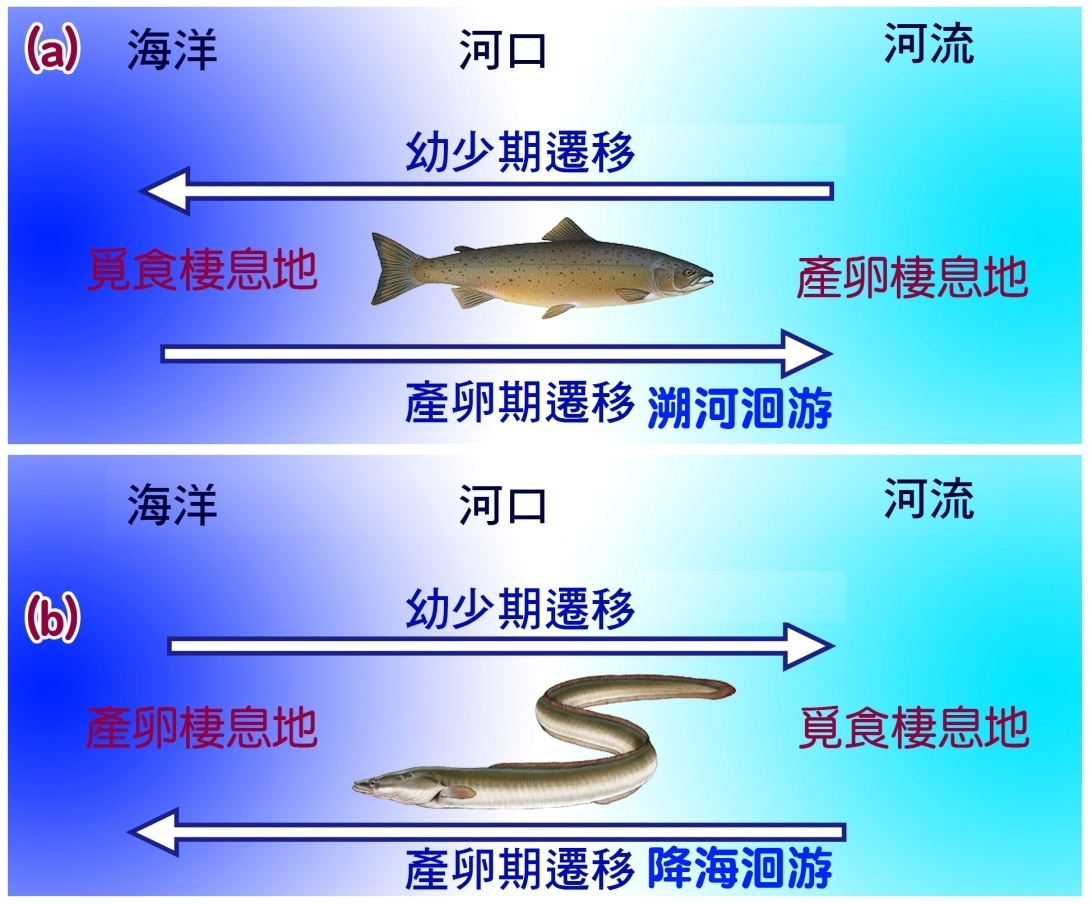
圖 5.2 ( a ) 溯河洄游 ( b ) 降海洄游 。
5.5 河流中的魚類活動
5.5.1 本土範圍和領土
河流對魚類活動有兩個重要特徵。首先,水流基本上是單向的,因此 上游運動,甚至僅僅停留在水流中,都取決於魚的主動遊動。相反,下游運動可以被動地進行,魚隨波逐流。在許多河流中,高瀑布或急流是上游運動的障礙。在屏 障上方的延伸段,只有當魚類表現出將下游巢穴減少到允許種群持續存在的水準的行為特徵時,種群才會持續存在 (Northcote, 1978) 。河岸和河底提供了一個相對穩定的地形,如有必要,魚類可以利用它來確定特定地點的方位。
在北溫帶河流的上游,鮭魚提供了與自然環境相關的社會組織的一個很好的例子。許多 鮭 科動物在水流湍急的小河中的礫石中產卵。雖然在 一些溯 河產卵的鮭魚中, 幼魚從礫石 中出來後很快就會下潛到海裡,但在其他物種中,在遷徙之前,溪流停留的時間較長 ( 在某些情況下,整個生命週期發生在淡水中 ) 。這些魚佔據了一個位置,使它們能夠接近漂流而過的獵物,同時也為它們提供了躲避水流的庇護所。 棲 地位點被視為領土。然而,一條魚會從一個 棲 地位點移到另一個 棲 地位點,當它離開一個 棲 地位點時,它會被另一條魚重新佔據。有些 棲 地位點比其他 棲 地位點更受歡迎,訪問這些 棲 地位點取決於給定位置的魚之間建立的支配等級 (Jenkins, 1969) 。這種社會制度被稱為部分屬地制。除了常住魚,也有更多的移動個體,當它們變得可用時,它們會從一個 棲 地位點移動到另一個 棲 地位點,但是很少或者沒有永久性的棲地位點連接。首選的地點可能是那些可以從漂流食物中獲得的能量和維持空間 棲 地位點的能源成本之間差異最大的地方 ( Fausch , 1984) 。這些位點可能成為鮭魚種間競爭的位點。
流過場地的水流保證了漂移有機物的持續供應。如果正在開發的資源沒有得到迅速更新,保護一個 棲 地位點可能不經濟。以生活 在基質中的 無脊椎動物為食的物種,或是以高度活動的獵物為食的物種,在覓食時往往分佈廣泛。魚可以作為個體生活在群聚或覓食中。即使是鮭魚,當它們變大並在其飲食中包括小魚,也更容易移動,不太可能保衛一個餵養 棲 地位點。有標記魚類的證據表明, 像擬鯉 這樣的鯉魚群可能佔據了一個家的範圍;然而,同樣的研究也表明,一部分種群是流動的,並不局限於一個家庭範圍 (Stott, 1967) 。這種可移動的部分可能是由從一個相鄰的群聚遷移到下一個群聚的個體,從而改變了它們的家園範圍。它也可能包括那些沒有 傾向於呆在 家園裡的人。
5.5.2 河流遷移
河流中的遷徙通常與繁殖、水位季節變化導致的覓食地開放或向越冬地區遷移有關。
河流的上游通常以含氧良好的水和足以防止泥沙 沉 降的水流為特徵。 食肉漁類 對魚卵和幼體的密度也可能較低。這一綜合因素可能是許多河流物種向上游遷徙產卵的原因。鮭魚提供了一個很好的例子,但其他淡水魚類也顯示出同樣的趨勢。在較大的河流系統中,這些上游遷徙覆蓋了很長的距離。生活在南美洲巴拉圭河系的大型 characins 從它們 在下查科的 覓食地遷移到 安第斯山麓 的繁殖地超過 400 公里 (Lowe McConnell, 1987) 。
在低緩河灘地河流中,季節性的高水位漫過河岸,淹沒了周圍的土地,為河魚開闢了廣闊的覓食區。亞馬遜系統中馬德拉河魚類的運動模式說明了這片被水淹沒的土地是如何被開發利用的 (Goulding, 1980) 。洪水初期,這些魚沿著馬德拉河的支流順流而下,在主河道產卵。產卵後,它們會向上游移動,然後散佈到森林底部覓食。當水開始下降時, 這些魚從森林 底部移到支流中,然後流到主要河流中。在枯水期,它們間歇性地向上游移動,然後隨著水位上升,再次向上游的支流移動。隨著魚的年齡增長,它們在河流系統中向 上游遊得越來越 遠,而下游的魚卵和 仔魚 補充了它們的數量,這些卵和幼體被沖到了主河道的下游。稍大的幼魚搬進泥沼、 洪泛平原 湖泊和被洪水淹沒的森林中,小而脆弱的體型迅速成長。
大多數河流系統的一個特點是幼魚會轉移到有遮蔽的淺水區,在那裡它們不太可能被沖走,並且有良好的食物供應。在河流系統的管理中,提供這些區域對於維持許多魚類種群至關重要 (Mills and Mann, 1985) 。
在溫帶和亞極地帶地區的河流中,大型幼魚和成魚可能遷徙到越冬地區。冬季,在英格蘭東部低窪的沼澤地,人們在與河流相連的船塢發現了高密度的魚類 ( (Jordan and Wortley, 1985) 。這些船提供魚類庇護所相當於自然懸掛的河岸或樹樁。
一些淡水物種在河流和湖泊之間遷徙。淡水鮭魚,包括褐 鱒 和 虹鱒魚 ,將在一條溪流中生活一年或數年,維持部分區域,並以漂流為食。然而,這些魚會向下 ( 或向上 ) 遷移到河系中的一個湖泊。在湖中,它們的活動範圍 要廣得多 ,因為它們不能再依賴不斷補充的漂流物。在性成熟後,它們會重新登上一條溪流產卵,通常會選擇它們孵化和度過生命最初幾年的溪流。類似的上游產卵遷移也出現在一些 熱帶湖棲物種 中,包括鯉魚 (Lowe McConnell, 1987) 。
5.6 湖泊運動與社會結構
在湖泊中,魚類的社會結構和運動與水流無關,而與沿岸帶和中上層帶的存在有關。前者類似於河流,因為存在固定的地形特徵。後者與公海相似,缺乏明顯的物理結構。
5.6.1 社會結構
生活在沿海地區的物種可能具有很強的地域性。綠色太陽魚就是一個例子,它基本上局限於北美東部的淺層、有植被的沿海地區 ( Gerking , 1959 年 ) 。其他的沿海物種可能生活在家裡的範圍內,例如湖棕 鱒 ,但這些範圍在空間上可能與北部梭魚一樣難以界定。在某些物種中,魚類基本上像梭子魚一樣獨居。在其他物種中,這種魚生活在較大或較小的群聚中,例如北美湖泊中的鮪魚、 非羅非魚 和鯉魚。像黃鱸這樣的物種在幼年時生活在群聚中,但隨著年齡和體型的增加,這種魚變得越來越孤獨 ( Helfman , 1981) 。
在大型湖泊中,海岸帶可能被分成幾個不同的棲息地,由不同的物種佔據。例如,馬拉維湖的西海岸線由交替分佈的沙子、雜草和岩石組成 ( McKaye 和 Gray, 1984 年 ) 。許多佔據岩石棲息地的慈 鯛 科物種 ( “ 姆布納 ” 物種 ) 表現出強烈的場地依附性,一生都停留在幾平方米以內。這種依附性可能是在馬拉維湖發現的許多慈 鯛 科魚類的原因之一,因為這種不願遷移的行為將確保佔據不同岩石海岸的種群之間幾乎沒有基因流動 (Fryer Iles, 1972 ; Lowe-McConnell, 1987) 。然而,有些人並不完全是久坐不動的。人工岩礁位於距離最近的天然 礁 1 公里處的砂質區域,被一些色彩鮮 艷 的非洲慈 鯛 “ Mbuna ” 物種所佔據 ( McKaye 和 Gray, 1984 年 ) 。這讓人想起了一個觀察結果,即河流種群包括常住種群和流動種群。
許多遠岸的 湖泊物種, 如棒狀 體和白魚生活在群聚中,這種在物理上不受限制的環境中的社會組織模式將在海洋 的遠岸物種 中再次遇到。
5.6.2 遷徙湖棲
一些湖棲魚類 在流入或流出的溪流中的遷移已經被提到。其他湖泊物種遷移到產卵地。例如,在美國的門多塔湖,白 鱸魚從湖 的表層帶遷移到湖邊的清澈的產卵區。
生活在中上層區域的一些種群的一個特點是垂直遷移,魚每天在水體中上下移動。這些遷移通常涉及以浮游動物為食的物種,如西非沃爾特湖的沙丁魚 clupeoid 物種 (Lowe McConnell, 1987) 。已經確認了兩種遷移模式 (Neilson 和 Perry, 1990) 。在 I 型遷徙中,魚在黃昏時上升,在黎明時下降。在 II 型遷徙中,下降是在黃昏,上升是在黎明。
I 型遷徙的一個例子是加拿大西部巴賓湖的紅 鮭 (Oncorhynchus nerka) 幼魚苗 (Brett, 1983) 。這些 幼 魚苗生活在以浮游動物為食的群聚中。夏天,當湖水分層時,魚在涼爽的深水中度過一天。黃昏時,它們會遷移到溫暖的地表水中覓食,當它們吃飽時會掉 進稍深 的水中。就在黎明前,它們再次在水面上覓食,然後又回到冰冷的深水中。這種垂直遷移的一個可能的作用是,它將魚苗被捕食的風險降到最低,同時仍然允許它們有時間在富含浮游動物的地表水域中覓食 (Clark and Levy, 1988) 。第二 個 ,但並非相互排斥的解釋是,幼魚苗通過在溫水中進食獲得生物能量優勢,在溫水中它們的新陳代謝率相對較高,但在冷水中休息時新陳代謝率較低 (Brett, 1983) 。最近對藍鰓太陽魚 ( Wildhaber and Crowder, 1990) 的一項實驗使後一種解釋變得更不可能。藍鰓鮪魚未能選擇溫度和食物水準的組合來最大限度地促進生長。湖泊中的 幼年湖擬鯉 表現為 Ⅰ 型和 Ⅱ 型遷移。
5.7 珊瑚礁和 岩岸 的社會結構和運動
珊瑚礁的物理複雜性提供了線索,這至少潛在地為魚類提供了重新找到避難所的方法。 許多礁魚 在珊瑚礁上的部分或全部生命中都表現出強烈的場所依附性。
生活在北美洲西部岩石海岸的石首魚 ( Oligocottus maculosus ) 等物種中,這種魚佔據了一個家的領地,但並不保衛自己的邊界。一隻 甲杜父魚 會在同一 個 岩石池 裡呆上 幾個星期或幾個月 ( Green 1971) 。其他物種具有很強的地域性。在珊瑚礁上,領土不僅可以抵禦同種物種,而且可以抵禦其他物種 (Sale 1980) 。這種種間領土防衛有時最強烈地針對那些與領地擁有者有相似飲食的物種 ( Myrberg Thresher, 1974) 。其他物種形成群聚,這些魚群允許魚類入侵領地,因為領地所有者被數量淹沒,例如,尖頭魚和鸚鵡魚的魚群會入侵雌魚的覓食區域 (Sale, 1980 年 ) 。其他淺礁魚類利用珊瑚礁上方的浮游動物。像咕嚕魚 ( Haemulon spp.) ) 這樣的物種沿著明確的路線從珊瑚礁遷徙到海草床上,返回珊瑚礁避難。
像一些石斑魚這樣的物種遷移到珊瑚礁的外緣產卵,這些魚在產卵地點形成大型產卵群。
珊瑚礁魚的特徵,在其他淺水 魚類區系中 ,魚類的數量較少,是白天活動的魚類群和夜間活動的魚類群在黎明和黃昏時的更替 ( Helfman , 1986) 。這種模式在時間上造成了魚類之間的隔離,這些魚類可能在捕食類似的獵物。
許多珊瑚礁魚類有能力回到它們的領地或棲息地,因此,如果它們被轉移或利用高水位離開它們通常生活的區域,它們可以返回家園。 斑紋 寡杜父魚 ,一種通常不到 150mm 長的魚,在被轉移到 90 米後,可以找到回到家裡的深水潭的路 ( Green, 1971) 。 鰕 虎魚,可以準確地從一個深水潭到另一個在低水位,以逃避干擾 ( Aronson, 1951) 。
當一個珊瑚礁區域的屬地魚類被移走後,它們的位置往往很快被其他人搶走 (Sale, 1980) 。這表明,至少有一部分棲息在礁石上的魚類是可移動的,一旦它們變得可用,它們就準備遷移到空曠的地方。
5.8 遠海魚類的社會結構與運動
5.8.1 遠海魚類
開闊水域的魚類可能是獨居的,但在許多物種中,個別魚類一生中的大部分或全部生活在群 游 裡。浮游 鯡 魚、鳳尾魚和其他棒狀體魚類、 鯖 魚等肉食性短鰭魚類以及以魚和魷魚為食的鮪魚,都是浮游魚類的好例子。遠海魚類生活在一個 沒有洞或裂縫 的環境中。它們的食物分佈在浮游植物或浮游動物的活動區,或者是利用 這些斑 塊的物種。在這種環境下,儘管魚類會聚集在漂浮的碎屑下面,或者會與近海鑽機等 結構物相關聯 ,但現場附著的可能性很小或根本不存在。
遠海魚類群的運動動力學還不為人所知。群 游 是隨意搬家,還是佔據一個分散的、不明確的家庭範圍?個體魚多久在群 游 之間交換一次,或者終身成為一個群 游 的會員?群 游 規模是否會因環境因素 ( 如食物供應 ) 而改變?
5.8.2 遠海魚類的遷徙
在它們的一生中,許多遠海魚類會進行跨越數百公里或數千公里的遠海遷徙。現在所知的大部分是大陸棚遠海魚類的遷徙,它們是或曾經是商業漁業的獵物。在北海 鯡 魚漁業的鼎盛時期,這種魚的遷徙反映在蘇格蘭東海岸和英格蘭北部的漁船和海岸工人上下遷徙,他們將魚掏空。
北海有幾種 鯡 魚在不同的礫石堤上產卵 ( 圖 5.3) 但 arden Jones, 1968) 。在多格河岸和英格蘭東海岸以西的河岸上有產卵地。這種魚在夏末從北海北部的覓食地向南遷移到秋季產卵的產卵地。這些 花魚向西北 方向遷徙到丹麥和挪威之間的斯卡格拉克 越冬地 。然後,在早春,它們向西北方向移動到飼養場,從而完成逆時針方向的 迴 圈。這種運動模式與北海的地表水流模式有關。 鯡魚幼魚被 帶到近岸,有遮蔽的海岸、海灣和河口充當 育雛區 ,其方式與河岸 幼魚利 用回水的方式相似。
遠海魚類的遷徙是眾所周知的。商業上重要的鮪魚物種表現出廣泛的活動 ( 圖 5.4) 。北美洲海岸外東太平洋的 北藍鰭 鮪魚在日本附近被重新捕獲,其他帶標籤的鮪魚則從另一個方向開始跨太平洋航行 ( Bayliff , 1980) 。
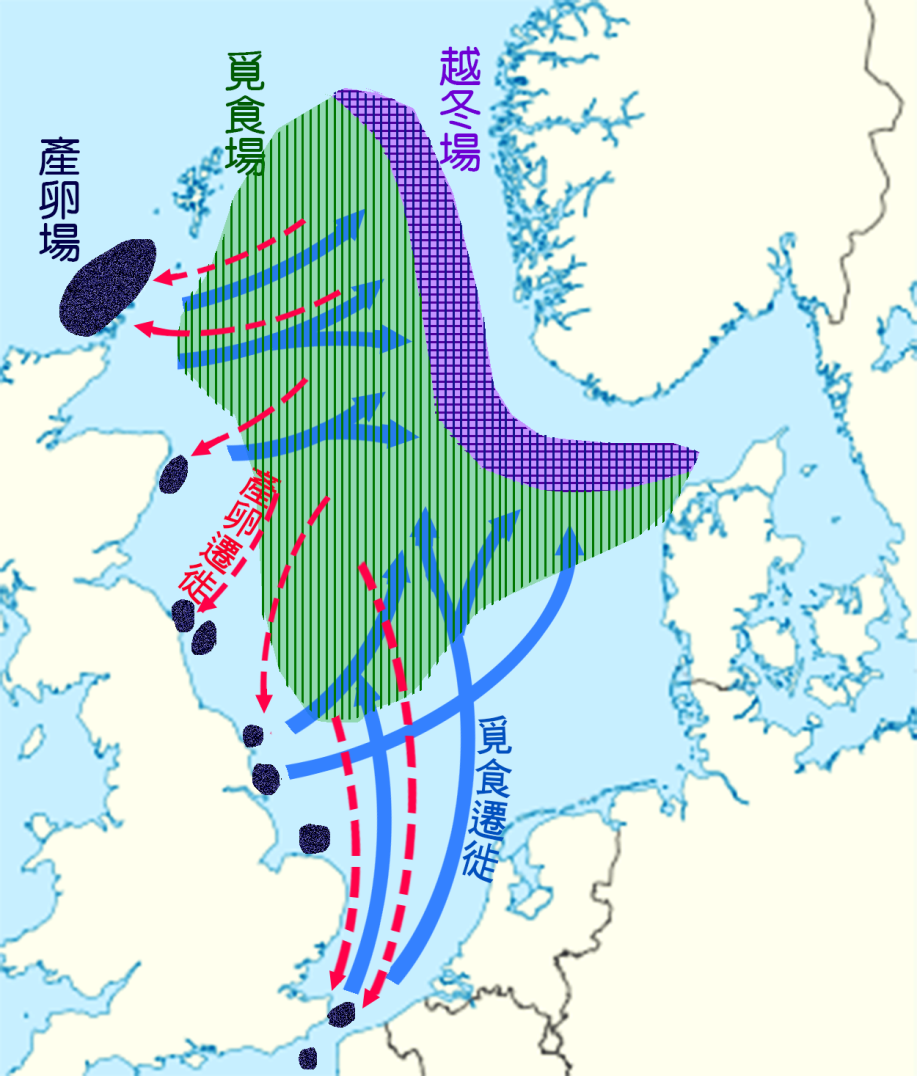
圖 5.3 北海 鯡 魚( Clupea harengus )的種群遷徙:產卵場為黑色。 覓食場,綠色直線區; 越冬場地,紫色 網格區 ; 產卵遷移,紅箭頭細線; 覓食遷移,藍箭頭粗線。 F.A.O. 1981 。
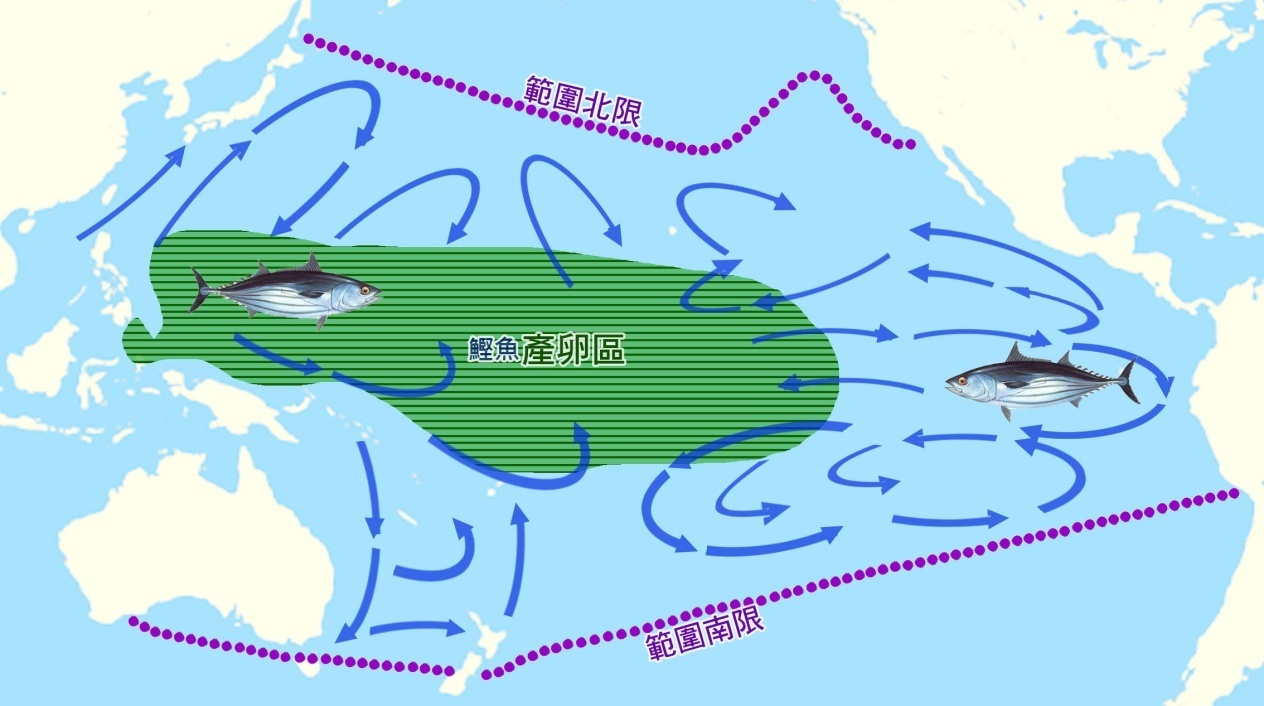
圖 5.4 鰹 魚在太平洋的遷移:產卵區 - 綠色水平線; 南北分佈極限 - 虛線。 F.A.O. 1981 。
5.8.3 遠海魚類的垂直遷移
一些遠海物種,如 鯡 魚,表現出與湖泊中遠海魚類相似的垂直遷移。在夜間, 鯡 魚會移到水面上,但白天它們會在較深的水域或接近底部的地方度過。許多燈籠魚 ( Myctophidae ) 在 幾百米深的 地方度過一天,那裡幾乎沒有陽光或沒有陽光。夜間,它們遷徙到表層,以 20 至 200 米深處的浮游動物為食。並不是所有的中層魚類都表現出如此大幅度的垂直遷移,有些魚類在每天的 週 期中都保持在相同的深度範圍內 (Marshall, 1979 ; Longhurst and Pauly, 1987) 。
對這種垂直遷移的適應性意義的理解將來自於對這種遷移的收益和成本的分析 (Clark and Levy, 1988 ; Neilsen 和 Perry, 1990) 。
5.8.4 底棲魚類的社會結構與運動
觀察生活在海底或靠近海底的魚類的困難意味著對這種物種的社會結構幾乎沒有詳細的瞭解。有標記的比目魚的運動表明,這些比目魚沒有屬地性 或家域行為 。在北海 南部, 浮游的幼體 鰈 被水流帶向荷蘭海岸。在蛻變期,幼魚棲息在有遮蔽的群聚上。隨著它們的生長,它們會移到更深的水裡。成年後, 鰈 魚會遷移到南方產卵,而用過的魚會返回北部的覓食地 (Harden Jones, 1968) 。在運動過程中, 海冰通過 一種稱為選擇性潮流輸送的行為過程利用潮流 (Arnold, 1981) 。在潮汐週期的靜水期接近尾聲時,魚離開 底部, 在水中停留數小時,在下 一個靜 水時返回底部。在中層水域,魚往往被主流潮流帶到下游。這種 魚遊得很活躍, 但速度很慢,很少超過每秒 1 個身長。游泳的方向並不總是與水流的方向一致。據估計,與不使用這種方法的魚相比,使用選擇性潮汐輸送的魚群可以節省 40% 的往返產卵場的能源成本。
以鱈魚為例的其他底棲魚類也表現出產卵、 覓食和越冬 棲息地之間的長距離遷徙。北極鱈魚從挪威北部水域遷徙到靠近挪威西海岸產卵。遠海的卵和幼體在沿海水域向北漂流,隨著成長中的幼魚移到靠近底部的地方開始它們的底棲生活。 底棲和遠 海魚類的產卵遷移導致卵 沉 積在盛行洋流通過被動漂流將發育中的卵和幼體帶向育嬰場的海域。
5.9 洄游
5.9.1 洄游分佈
所有或部分魚類定期在淡水和海水之間移動的物種包括一些最著名的洄游物種,包括北太平洋的太平洋鮭魚、歐洲和美洲鰻魚以及希爾 薩 ( 印度和東南亞的 hilsa ilishaj ) 。 溯 河魚類包括原始但特殊的無顎七 鰓鰻 ,這種生活方式在原始 硬骨目、棍狀目、鰻形 目和 鮭 形目中得到了很好的體現。它在進化更高級的硬骨魚中更為零星地出現 (McDowall, 1988) 。
溯河產卵魚類和水下產卵魚類的分佈存在地理格局 (Gross, 1987 年; McDowall, 1988 年 ) 。在 30 ° 到 60 ° 北緯 之間, 溯河產卵物種的頻率最高 ( 圖 5.5) 。這一高峰反映了七 鰓鰻和 鮭魚的地理分佈。在大約 60 ° 北緯和 50 ° 南緯之間發現 了降海物種 ,在熱帶和亞熱帶有 一個峰 值 ( 圖 5.6) 。這反映了鰻魚 ( 鰻鱺科 ) 和 鯔 魚 ( 鯔 科 ) 的地理分佈。
如果用溯河 物 種的比例來表示,則地理格局更加清晰。隨著緯度的增加,溯河產卵種類的比例增加,而下降的種類比例隨著緯度的增加而減少。有兩種解釋可以解釋這些分佈,而 不是相互排斥的。這種模式可能反映了這些遷移類型的系統發育相關性。鮭魚是冷水魚,也碰巧是溯河產卵魚類,而鰻魚基本上是來自溫暖水域的一個群體,也碰巧 是降海產卵 魚類。根據這一解釋,緯度與遷移類型之間的相關性是巧合的。
一個更有趣的可能性是,這種模式反映了淡水和不同緯度海水的相對生產力 (Gross, 1987) ,這一假設表明,在高緯度地區,海洋的生產力高於淡水。從淡水遷移到海洋的魚會遷移到更肥沃的覓食地,因此與呆在淡水中的魚相比,它們的生長速度會有所增加。在低緯度地區,淡水往往比海洋更有生產力,而遷移到更肥沃的覓食地會 把魚從海水 中帶到淡水中。與生產力梯度有關的任何遷移都可能受到一些因素的影響,例如需要避免極冷的海水或淡水棲息地可能乾涸的乾旱時期。
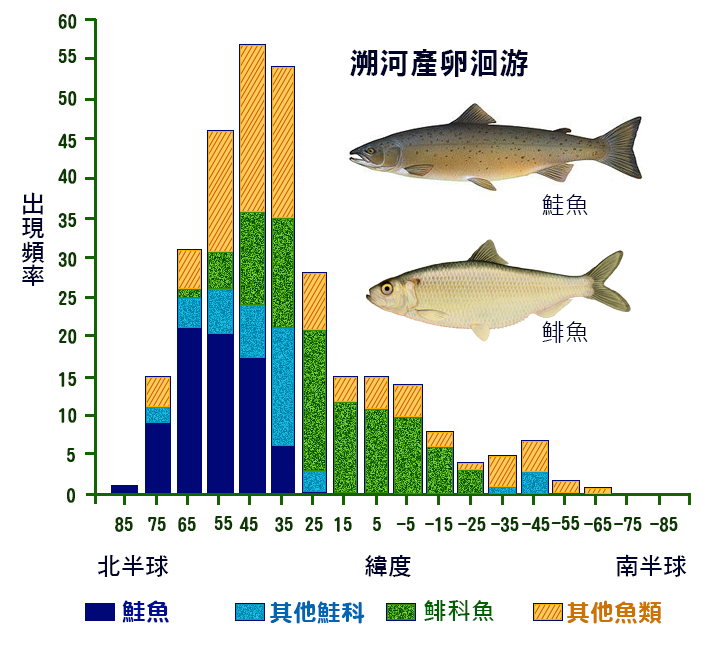
圖 5.5 溯河 洄游物種相對於緯度的頻率。 向北緯度顯示為正值,向南緯度顯示為負值。 重點:( 1 )鮭魚;( 2 )其他 鮭 科魚類;( 3 ) 鯡 形目 ;( 4 )其他魚類。 McDowell1988 。
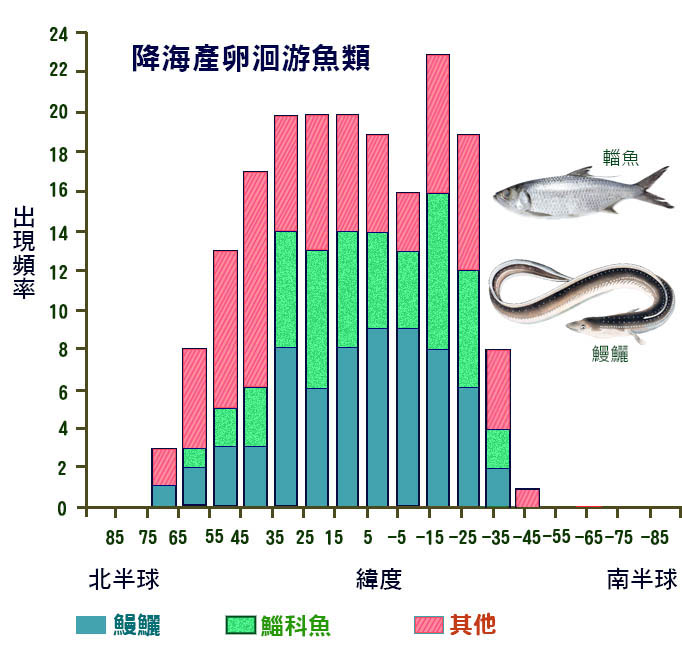
圖 5.6 降海洄游 物種相對於緯度的頻率。 關鍵:( 1 )鰻鱺屬 ; ( 2 ) 鯔 科 ; ( 3 )其他魚類。 McDowell1988 。
5.9.2 溯河魚類的社會結構
在一個生命週期內可能表現出的社會結構的範圍 可以從溯河 產卵鮭魚 ( 如大西洋鮭魚或北太平洋的紅 鮭 ) 中得到很好的說明 (Mills, 1981 ; Foester , 1968) 。在卵產卵的紅礫石層中出現後,幼小的大西洋鮭魚佔據了一個覓食區域,這為它提供了一系列可以覓食的地方。一年或一年以上後,它會轉變為幼 鮭 ,並向下游的海面移動,幼 鮭 在群聚中向下游移動。一旦進入大海,這些魚就會遷移到格陵蘭島沿岸的覓食地。最終,成熟的成蟲會遷移回淡水中,通常回到它們出生的溪流。
在繁殖場上,雄性保衛雌性在其中挖掘產卵的紅色區域。在這些繁殖區域,成魚不進食。一般的模式 與刺魚 相似,只是在孵化後,幼魚會向下游移動到湖中。在湖裡,它們生活在以浮游動物為食的群聚裡。通常在 湖裡呆上一 年後,它們會窒息並在群聚中遷徙到海裡。它們穿過北太平洋,基本上是沿著北太平洋逆時針方向的 大旋回 運動。隨著成熟期的臨近,它們又回到淡水中。同樣,在產卵地上,雄性保衛雌性產卵的領地。
在大西洋鮭魚和紅眼鮭魚中,有些魚不遷徙,而是在淡水中度過一生。在大西洋鮭魚中,這些是典型的雄性,它們在很小的 年紀就性成熟 ,體型也很小。在大 馬哈魚 ( 紅 鮭 、藍背 鮭 ) 中,雌魚和 雄鮭魚 都可以呆在淡水中。它們被稱為 銀鮭魚 - 青幼鮭魚 ( Kokanee) 。
5.10 返鄉
有很好的證據表明,許多遷徙物種為了產卵而返回或靠近出生地。在一些洄游鮭魚種群中,只有不到 5% 的回 游 魚類未能返回其出生的溪流 (Quinn, 1984) 。在北海的 鯡 魚和 鰈 魚中,來自不同種群的成魚在不同程度上可能會在覓食地上進行不同程度的混合,但它們的產卵遷移會將它們帶回出生產卵地。魚是如何回到它們產卵的繁殖地或回到它們的家鄉池塘的?
5.10.1 返鄉機制
環境提供了許多線索,理論上,這些線索可以被歸巢的魚使用 (Smith, 1985) 。這些因素包括水流、溫度和鹽度的梯度、嗅覺刺激、白天太陽或夜晚 恒 星的位置、白天天空中偏振光的模式以及地球磁場的變化。這種過多的潛在線索提出了兩個問題:哪些線索可以使用,哪些線索在遷移過程中使用?
實驗研究表明,一些遷徙物種原則上可以使用這些線索中的一些,但問題仍然存在 — 它們是否使用這些線索?鮭魚提供了很多證據 ( 也有很多爭議 ) 。
人們普遍認為,一旦遷徙的成年鮭魚到達淡水中,它就依賴嗅覺。找到合適的產卵地。鼻孔堵塞或嗅覺神經受損的鮭魚無法區分 “ 家 ” 的水和其他水 (Hasler 和 Scholz, 1983) 。對於所使用的嗅覺線索的性質存在爭議。有一種假設認為,幼魚在淡水中的幼魚在幼年期就學會了家鄉溪流的嗅覺特徵。當它在海裡覓食的時候,它會保留這種記憶,作為一個回歸的成年人,它會跟著它的家鄉溪流的氣味 游 向上游 (Hasler 和 Scholz, 1983) 。另一種假設認為,嗅覺線索是由幼魚在原流中釋放的資訊素提供的,而返回的成年魚對其種群特有的資訊素具有天生的反應能力 ( Stabell , 1984) 。甚至有可能魚使用兩種機制的組合,充分利用所有的嗅覺線索。
在長時間的海洋遷徙之後,這些魚是如何找到它們的家鄉河口的,還有很多不確定性。一種可能性是,這些魚表現出輕微的方向性運動,將它們帶到適當的海岸線 ( 有偏差的隨機搜索 ) ,然後在海岸線上搜索攜帶 適 當家的水氣味的 淡水羽流 。另一個極端是這種魚有可能擁有發達的導航能力,這使它們能夠直接和高精度地回家。導航能力假設是基於對太陽的位置和位置變化的探測,以及在陰天的情況下,地球磁場。其他研究表明,即使是在它們遷徙的海洋階段,鮭魚也在使用嗅覺信號,與鹽度和溫度的電流和 梯度 有關。 Quinn(1984) 總結了太平洋鮭魚洄游回淡水的海洋階段是一種定向而非有偏的隨機運動的證據。證據包括以下觀察結果: ( i ) 在海洋中某一特定地點標記的魚分散到廣泛的母系溪流; (ii) 標記研究表明,從界定廣泛的飼養區向精確的沿海地區移動的速度相對較快; (iii) 一個種群中的個體以顯著的時間聚集在它們的家鄉河口; (iv) 遷移鮭魚在它們標記的地點到它們重新捕獲的地點之間移動的速度表明它們正在積極地定向。為了對遷徙魚類的精確定位有一個令人愉快的懷疑的觀點, Leggett(1984) 的一章應該閱讀。
5.11 捕撈的影響
許多商業上最重要的物種是洄游魚類。 1978 年世界漁業產量最高的 25 種海洋物種中,有 24 種是洄游物種 (Harden-Jones, 1981 年 ) 。在歐洲人到來之前,太平洋鮭魚是西北太平洋美國人的主要食物。隨後,同一物種成為主要漁業和相關罐頭工業的目標。大西洋 鯡 魚和鱈魚都是遷徙物種,它們都支持歐洲主要的工業化漁業 (Cushing, 1988) 。
許多遷徙物種的一個特點是,繁殖發生在空間受限的繁殖地,例如鮭魚所用的源頭和大西洋 鯡魚所 用 的礫石堤,然而,特定產卵產生的魚的重量遠遠大於這些產卵場上或附近所能容納的魚的重量。這種放大作用取決於魚在某些情況下長距離移動到其他更豐富、更大 的覓食區。與非洄游魚類相比,洄游魚類在覓食地的增長速度可能是洄游習性進化的一個關鍵因素。這一增長率的增加將使它們能夠迅速地穿過最容易被捕食的體 型。這將使他們能夠獲得更 大 和更有利可圖的食物。體型較大的雌性更多產,體型增大可能會使雄性在 雄性間的配偶 競爭中佔優勢。
然而,這種洄游習性可能使它們容易受到密集捕撈的影響。以鮭魚為 例,返回的魚類聚集在鄰近其家鄉溪流的局部沿海地區。許多洄游魚類在容易被商業漁船用電子方法探測到的群聚中移動,一旦被發現,魚群就會向漁具提供高密度 的魚。遷徙也可能使魚類通過不同國家或經濟集團管制的捕魚區,這將使漁業管理更加困難,因為有更多的利益需要滿足。