
魚類生態學
1. 水環境與魚分類
2. 魚類生理學
3. 環境條件對魚種分佈的影響
4. 生物因數與魚類群落結構
5. 移棲地 與魚群行動
6. 攝食與生長
6.1 概述
6.2 河流環境中的攝食生態
6.2.1 湍急溪流魚類的攝食生態
6.2.2 低地緩流魚類的攝食生態
6.3 湖泊攝食生態
6.4 河口的攝食生態
6.5 海洋魚類的攝食生態學
6.6.1 岩石和珊瑚礁魚類的餵養
6.6.2 大陸棚的攝食生態學
6.6.3 遠海覓食生態
6.6.4 南極魚類的攝食生態
6.6 食品的檢測和選擇
6.7.1 硬骨魚和 食草 動物的檢測和選擇
6.7.2 食肉魚類的檢測與選擇
6.7.3 最佳覓食
6.7 攝食生態形態學
6.8.1 篩檢程式 饋 線
6.8.2 腐食魚類
6.8.3 食草魚類
6.8.4 食浮游魚類
6.8.5 食軟體動物魚類
6.8.6 食肉魚類
6.8.7 生態形態學假說
6.8 魚類的營養種類
6.9 食物利用
6.9.1 能源收支
6.9.2 營養素收支
6.10 食物消耗率
6.11 魚類增長
6.11.1 魚類體型與棲息地的關係
6.11.2 食物和環境特性對生長的影響
6.11.3 重量與長度的關係
6.11.4 生長測量
6.11.5 魚的老化
7. 生活史和種群動態
8. 魚類應用生態學
第六章 攝食與生長
6.1 概述
魚必須從食物中獲得大量和微量營養素 ( Halver , 1989) 。大量的營養素是蛋白質、 脂類和 少量的碳水化合物。 這些宏量營養素 提供基本的組成部分,即蛋白質中的氨基酸、 脂類中的 脂肪酸和碳水化合物中的醣類,它們被用來修復身體組織的損傷,並合成新的肌肉。大量營養物質也是在呼吸過程中被氧化的燃料,產生能量。這種能量被用來維持一個功能正常的身體,游泳和合成身體生長或生殖產品 ( 卵子或精子 ) 形式的新組織。微量營養素是必需的維生素和礦物質,少量的,是有效新陳代謝所必需的。微量營養素和微量營養素的需求可能從許多方面得到滿足。在硬骨魚中,廣泛的潛在食物反映在廣泛的飲食和餵養方法上。銀 鱈 和軟骨魚在其攝食生態上的多樣性要少得多。
潛在的食物種類在很大程度上反映了自然環境。一個物種的活動範圍取決於它探測、獲取和加工食物的能力。綠色植物光合作用的初級生產是魚類最終食用的幾乎所有宏觀營養素和有機微量營養素的最初來源。然而,與陸生環境相比, 食草性 脊椎動物常常是動物群的顯著組成部分,而 食草性 魚類通常不是水生食物網的主要組成部分。其中一個原因在於水生環境的基本特性。水環境中的大部分初級生產是由單細胞藻類產生的,它們通過水的浮力維持在水體的發光區域。這些藻類很小,通常 2-200 微米 ( μ m ) 大小。需要一個細 網或篩檢 程式來收集它們。一旦收穫,細胞必須被打破。通常情況下,魚類通過 食肉性無脊椎動物 或其他無脊椎動物和脊椎動物食肉動物間接利用這些藻類。 錨定植物 ,包括藻類和開花植物,僅限於河流、湖泊和海洋邊緣的淺水區。即使在這裡,被水流或潮水沖刷的不 穩定基質通常 也會阻止植物生根。 食草動物 是魚類群落的重要組成部分,在那裡有堅硬 的基質 ,被硬殼藻類佔據,或者在大型植被可以生存的淺水區,或者在浮游植物達到高濃度的高度肥沃水域。
以腐爛物質為基礎的食物網及其相關的細菌、真菌和其他微生物群也包括魚類。雖然有些魚類可以以腐爛的物質和有機碎屑為食,但大多數魚類直接或間接地以碎屑無脊椎動物為食。腐爛物質的原始來源可能在群落 內部, 其來源是本地的,也可能是起源於另一個群落但已進入水生群落的物質。這種物質被稱為外來物。因此,湖濱的睡蓮腐爛的葉子、莖和根形成了一個本地的碎屑來源,但是從邊緣樹木掉入湖中的葉子是一種外來的營養和能量來源,任何陸地節肢動物或其他落入水中的動物也是如此。
在討論魚類的攝食生態學時,一個有用的術語是 guild行會- 同功群 ( Begon 等人 , 1989) 。在同一棲息地中以相同營養水準為食,以相似的方式開發同一類資源的魚類,構成了一個行會 ( 同功群 ) 。
6.2 河流環境中的攝食生態
6.2.1 陡急溪流的攝食生態學
在水源地,經常發生的急流阻礙了有根植物的生長和大量浮游植物的生長。初級生產是由藻類和苔蘚產生的,這些苔蘚包裹著河床中的石頭和岩石。隨著分 汊 池結構的發展,溪流魚類食性的多樣性增加。
美國肯塔基州東部的一條溪流說明了這一進程 ( Lotrich , 1973) 。只有一種鯉科動物, Semotilus atromaculatus ,生活在 最 頂端。它以落入溪流中的陸生無脊椎動物為食,而不是溪流中產生的食物。隨著河流秩序的增加,魚類的數量也在增加。第一批是以水生昆蟲為食的,其中一個物種,石首魚 ( Campostoma anomalum ) 以生長在溪流中的藻類為食。它們都是本地的食物來源。隨著溪流的進一步擴張,又出現了兩個餵食 同功群 。一個由三個物種組成的群體,都是中心 生殖綱 ,以水生脊椎動物為食,而鯉魚 ( Pimephales notatus ) 的胃中有碎屑。食性動物群落的生物量和豐度是它們在河流群落中相對重要性的第一個近似值 ( 表 6.1) 。在伊利諾州的一條溫暖的水流中也可以看到類似的模式 (Schlosser, 1982) 。淺層、不穩定的棲息地被 一般食蟲動物 佔據,而在較深和較穩定的池塘棲息地中,主要的食性同 功群是食蟲 食肉動物和底 棲食蟲 動物。
下游的本地食物越來越重要的一種模式也表現在一條熱帶小河的魚群中,即馬來西亞的 Sungai-Gombak(Bishop, 1973) 。在小源頭支流中, 斑腹魚以 陸生昆蟲、蜘蛛和水生昆蟲為食。另外兩個物種是雜食性的。再往下游,出現了其他食性 同功群 ,包括以水生無脊椎動物和脊椎動物為食的食肉動物。雖然本土食物的貢獻增加了,但主要的能源供應來自外來植物材料,在較小程度上,陸生無脊椎動物。
河畔大樹冠植被遮蔭量對溪流魚類的攝食生態有重要影響。在溪流被濃蔭遮蔽的地方,溪流中幾乎沒有初級生產力,主要的能量輸入是從陸地植被和陸地無脊椎動物身上掉落的樹葉、花朵、水果和其他碎屑,它們被困在表層薄膜中。在這些溪流中,主要的食性 同功群是 雜食性動物,它們既可以吃動植物,也可以捕食陸生或水生無脊椎動物。在巴拿馬的小河中,藻類生長在河床上,沒有遮蔭。這種藻類被主要由長鰭 鯰 魚組成的食藻動物同 功群吃 掉 ( Angermeier and Karr, 1984 ; Power, 1987) 。在巴拿馬溪流中,藻類飼料和以陸生植物為食的物種的生物量隨著溪流的大小而增加,而以水生無脊椎動物為食的物種的生物量則下降。
大多數溪流魚類的一個特點是它們能夠攝取許多不同種類的食物,因此它們能夠對食物供應的變化作出反應。這種靈活性可能在不同的河流環境中以不同的方式表現出來。在巴拿馬的一條溪流中,在旱季,當食物資源減少, 魚被擠 到禁區時,它們的飲食變得不那麼相似 ( Zaret 和 Rand, 1971) 。相比之下,在南美洲北部的溪流中,由於更多物種的首選食物減少,更多的物種 轉而攝入 碎屑,因此它們的飲食在旱季變得更加相似 (Lowe McConnell, 1987) 。溪流魚類在攝食方面的這種靈活性反映在它們缺乏高度專業化的形態或生理適應能力。 Winemiller (1990) 對熱帶淡水溪流中食物網的複雜性進行了描述和定量分析。
表 6.1 由其定義的魚類攝食 同功群的 豐度和總生物量估算, 肯塔基州東部 一、二、三級序 溪流的典型飲食。 Dat Lotrich (1973).
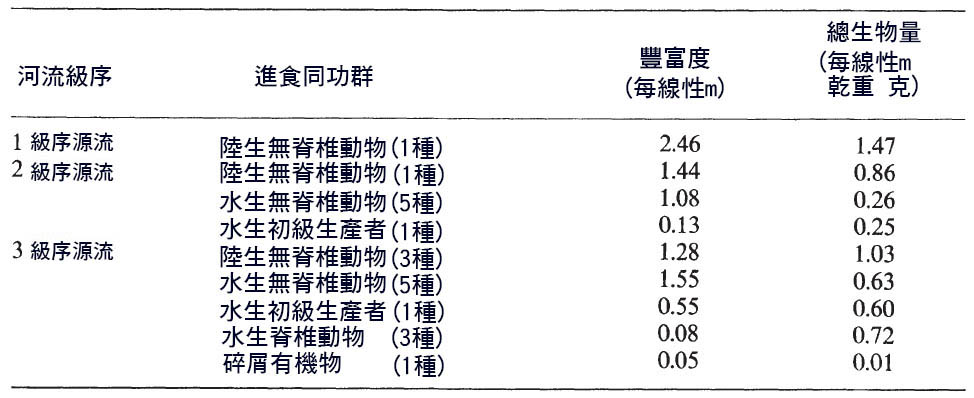
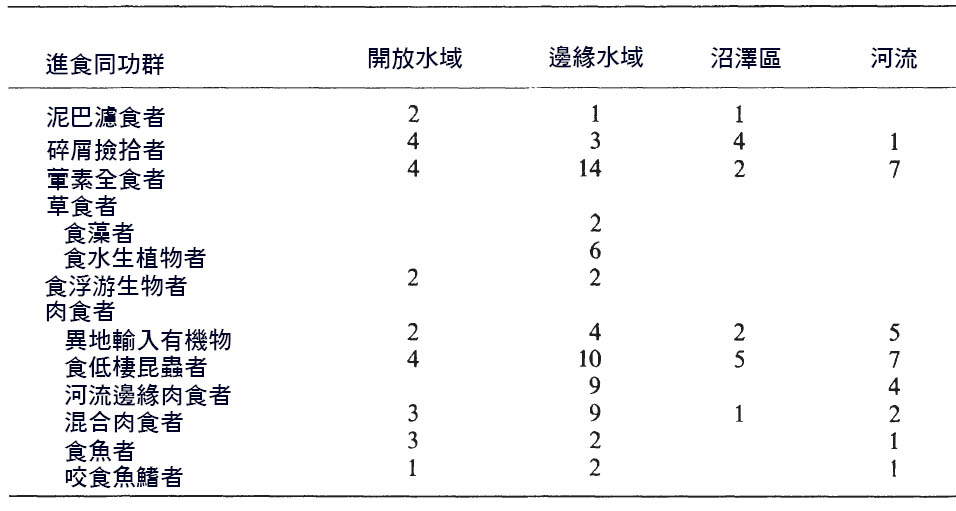
表 6.2 非洲薩伊河棲息地中的食性 類群屬數 Data from Lowe-McConnell1987)
6.2.2 漫灘河流的攝食生態
河灘地棲息地多樣性反映在食性群落的多樣性上。 Lowe McConnell(1987) 對中部非洲薩伊河的食性 同功群進行 了分類 ( 表 6.2) 。兩個棲息地包含了大多數的 同功群和 大多數魚類物種,是河道的沿岸地帶和海灣和沼澤地。西非尼日爾河也描述了類似的 同功群 ( Welcomme , 1986) 。
當這些河流氾濫時,被淹沒的地區提供了潛在的豐富的食物來源。淹死的陸生無脊椎動物成為 食肉動物和雜食 動物的食物,土壤中的營養物質可以支援浮游植物和相關的浮游動物的大量繁殖。當洪水淹沒的地區是森林時,樹木會再次收穫 ( 圖 6.1) 。亞馬遜河流域的許多魚類移到森林的地面上,以水果為食,種類包括 像巨腹巨腹魚 這樣的大型魚類。對於這些物種來說,森林洪水時期是主要的攝食季節,而在旱季,當河流被限制在主河道內時,它們的胃可能基本上是空的,魚依賴於洪水期間釋放的脂肪儲備。
在一些熱帶河流的 魚類區系生物 量中,碎屑魚類占很大比例 (Bowen, 1984) 。在南美洲的大河中, 鯪 脂鯉科 和 無齒脂鯉 科 的特徵科非常豐富,它們利用 森林被淹地面 和河灘上水 窪 池塘中的碎屑。
淹水區、河灣和回水區是河流魚類幼魚的重要育嬰區。滯流的水域使豐富的動植物群得以發展,微生物適合作為剛孵化幼體和 幼魚 的食物 (Mills 和 Mann, 1985) 。
溫帶河流中魚類的 食性類群不如 熱帶河流中的多樣性。 動物區系往往 以雜食動物為主,如 湖擬鯉 、一般食肉魚類,如鱸魚、梭子魚 ( 圖 3.6) 。雖然 將河漫 灘魚類劃分為 食性類群是 方便的,但它們中的許多仍保留著食物的靈活性,這是源頭和溪流動物群的一個特點。
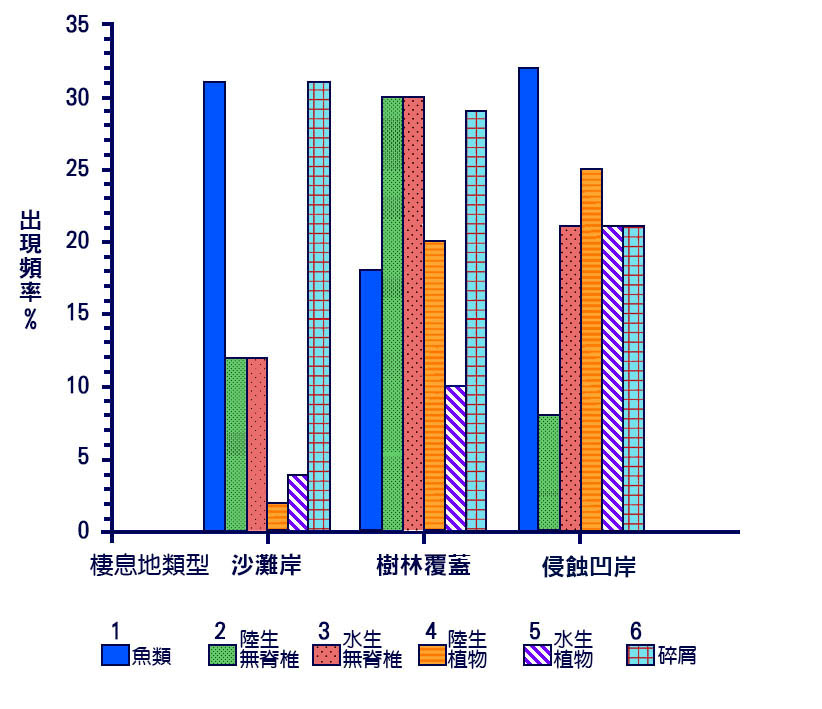
圖 6.1 南美洲里約熱內格羅河河灘地的三個常見棲息地中的飼養公會。棲息地有河灘、洪水氾濫的森林和木本海岸。頻率是指在每一種類上顯著進食的物種的百分比。食品類別關鍵:( 1 )魚類;( 2 )陸生無脊椎動物;( 3 )水生無脊椎動物;( 4 )陸生植物資料;( 5 )水生植物資料;( 6 )碎屑。 Data from Goulding et al. (1988).
6.3 湖泊攝食生態
陸生外來物質的重要性隨湖泊面積、形狀和海岸線的複雜性而變化。在小湖泊中,這些投入物可能為食物網提供重要的基礎 (Hanlon, 1981) ,但在大型湖泊中,原生初級生產將占主導地位。
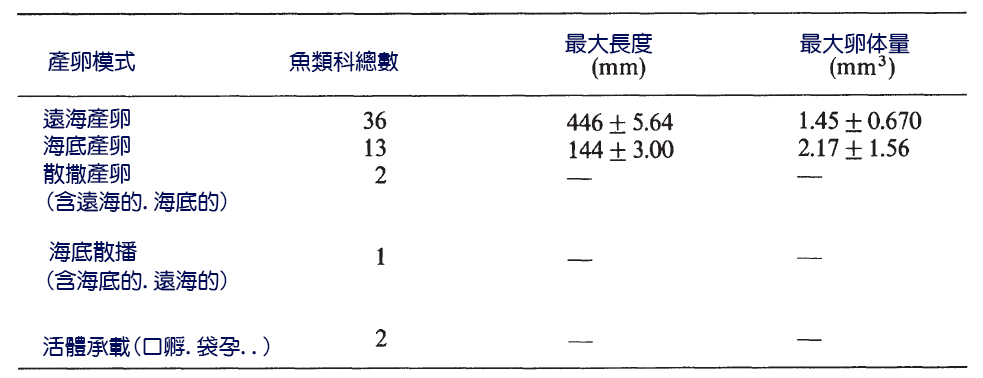
非洲五大湖、馬拉威湖、坦桑尼亞湖和維多利亞湖的特點是其慈 鯛 科魚類群落的物種多樣性很高。這種多樣性與攝食生態學中廣泛的適應性輻射有關。 Witte(1984) 在維多利亞湖姆萬扎灣收集到的亞 科麗魚 ( 雜金色慈 鯛 ) 中發現了大約 11 種營養類型。這種豐富的動物群隨後被引入的食肉動物尼羅河鱸魚 (Lates niloticus ) 嚴重耗盡。食蝸牛類軟體動物,按捕食蝸牛的方式可分為 三個亞類 。一些物種抓住了蝸牛的柔軟部分,然後將其 縮回殼 中,取出蝸牛。有些人用 牙齒咬碎貝殼 。其他魚種則在咽喉 部的咽骨 之間擠壓貝殼 ( 圖 6.2) 。確定了褐藻的 兩個亞類 。一組從植物中刮取藻類 ( 附 生刮板 ) ,另一組從石頭上刮取藻類 ( 表層刮片 ) 。食肉動物包括食昆蟲魚類、食浮游物魚類、 吃蝦魚類 、吃螃蟹魚類和從其他魚類身上採集寄生蟲的清潔工魚。
食魚的樸麗魚 ( 慈 鯛 科 ) 又分為 三個亞類 。一組以整條魚為食,有些通常 吃非慈鯛 科魚類,另一些則以慈 鯛 科魚類為食。第二組以其他慈 鯛 的卵和魚苗為食。這些專吃幼魚體的魚有各種各樣的技術, 可以使口中 孵育的母魚吐出它們 的幼仔 ,然後搶吞食。第三組以其他魚類 側面刮 下的鱗片為食。其他種類為碎屑動物、浮游植物和大型植物。在墨西哥灣的拖網樣本中,捕獲的主要 是食蟲魚類 、食浮游物魚類、食碎屑和浮游植物魚類。雖然這一營養類物種的數量最多,但食魚類占總漁獲量的不到 1% 。雖然在許多情況下,一個物種的嘴的位置和形狀以及牙齒的排列可能與其通常的食物有關,但慈 鯛 仍然保持著營養上的靈活性,如果它們變得豐富,它們就會轉而食用其他食物。
在坦桑尼亞湖,有一些特有的 鯡 魚類物種 (clupeoids) ,它們在幼年時以浮游植物和浮游動物為食,成年時以浮游動物為食。其中一個物種, Limnothrisa miodon ,在成年時有一個更普遍的飲食,吃昆蟲、蝦和夜蛾的幼蟲 (Lowe McConnell, 1987) 。
在溫帶湖泊中,可以識別出類似的營養類型,但已經注意到的許多熱帶物種在覓食方面的靈活性更為明顯。這往往模糊了類別之間的區別 ( Keast 和 Webb, 1966 ; Keast , 1978) 。在安大略省的 Opinicon 湖,漂浮水面的浮游動物被 溪銀漢魚 吃掉,但這也帶走了水面上的昆蟲。有兩種太陽魚, 藍鰓魚和 駝背太陽魚 ,都是在海濱覓食的食肉動物。大 駝背太陽魚 可以吃蝸牛,用 它的咽骨壓碎 蝸牛殼,而 藍鰓魚以 昆蟲幼體浮游動物為食。食 肉岩鱸 、 龍眼鱸和黃 鱸隨著體型的增長,所占比例越來越大。 兩種食 魚動物,北梭魚和大口鱸魚,在生命的最初幾周變成了食魚動物。底棲牛頭魚、真 鮰 Ictalurus ,晚上以底棲無脊椎動物為食。可能沒有反映強烈季節性特徵的溫帶湖泊。冬季大型植物死亡,浮游植物密度呈現明顯的季節變化。
在北美洲的五大湖中,一種 叫 灰西鯡 的大西洋 鯡魚 專 吃浮游物,在 20 世紀 60 年代變得極其豐富,並成為主要漁獲。 20 世紀從東海岸入侵五大湖的食浮游物魚類數量激增,可能是由於沉重的捕魚壓力和七 鰓鰻的 捕食,及本土 食魚性魚類 和浮游動物數量大幅度減少的結果 (Smith, 1968 ; Christie, 1974) 。 圖 6.3 顯示了美國大湖的簡化食物網。
在更偏北 的湖泊中,吃水面浮游動物的角色往往由白鮭魚物種 (Coregonus spp.) 承擔,而一般食肉動物的角色則由鮭魚和紅點 鮭 (Salmo 、 Salvelinus) 承擔 ( Svardson , 1976) 。沒有任何物種佔據軟體動物、草食動物或碎屑動物的營養類別,但這些食物類別中的每一種都可能被一系列物種作為更廣泛飲食的一部分攝取。
圖 6.2 非洲維多利亞湖的 麗魚慈鯛的咽骨 顯示牙齒形狀和食物之間的關係:( a )食碎屑魚;( b ) 食蟲魚 ;( c )食魚魚類;( d )食無脊椎 / 肉食魚;( e )食無脊椎。 Greenwood (1984).
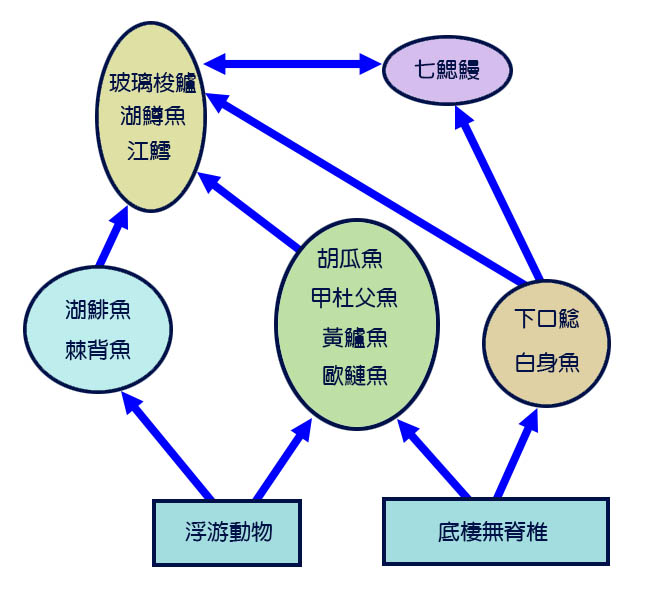
圖 6.3 北美蘇必略湖的簡化食物網。 Cohen and Stone (1987).
6.4 河口的攝食生態
對於那些能夠忍受苛刻的身體條件的動物來說,河口提供了高產的飼養場。一種主要的能源是進入河口的碎屑,或是從河口植物腐爛而來的碎屑,如鹽 沼 植物、海草和紅樹林。魚類 主要以底棲 動物 ( 小型底棲動物和大型動物 ) 中或與其密切相關的無脊椎動物為食。小型底棲動物是一種無脊椎動物, 成年體尺小於 45 微克 ( μ g) 乾重 ,通常用於長度小於 60 mm 的小魚 (Gee, 1989 年 ) 。最常見的獵物是原形 橈足類 ( 甲殼綱 ) ,它們的活動可能使它們比小型底棲動物的其他組成部分更容易受到魚類的攻擊。隨著魚類的生長,它們使用大型動物群的組成部分,如多毛類蠕蟲、大型甲殼類動物、軟體動物和小魚。幼年比目魚會切下 雙殼類的 虹吸管和羽扇蟲的尾巴 ( Trevallion , 1970) 。
碎屑 ( 及其相關的動植物群 ) 也被河口和沿海鹽 沼 中的一些魚類所利用。物種利用這種豐富但品質差的食物來源的有效性各不相同。包括 鯔 魚在內的 鯔 魚以碎屑為食 ( Odum , 1970) ,儘管幼年 鯔 魚可能不如成年 鯔魚更 好地適應利用這種資源 (White, 1986) 。加拿大底 鏘 魚雖然全年都生活在鹽 沼 中,在食物短缺的時候會攝入碎屑,但體重減輕,以碎屑為食的死亡率很高 (White, 1986) 。
6.5 海洋魚類的攝食生態學
6.5.1 以岩礁岸和珊瑚礁魚類為食
這些淺水海洋棲息地結構複雜,為魚類提供了多種謀生方式,這反映 在礁魚的 營養生態中。最值得注意的是,食草魚類可以構成 魚類區系的 重要組成部分。
在珊瑚礁上的魚類組合中可以識別出一系列的食性 同功群 (Safe, 1980 年; Longhurst 和 Pauly, 1987 年 ) 。有些物種以珊瑚礁表面或接近其表面為食。另一些魚類利用珊瑚作為庇護所,但在水體或遠離礁石的棲息地 ( 包括相鄰的海草床 ) 中覓食。珊瑚礁群落的食物網主要包括水體中浮游植物的初級生產、 包殼藻類 的初級生產以及來自動植物來源的珊瑚和碎屑的藻類共生體。 包殼藻被幾種食草 動物群所利用,包括 雀 鯛 ( 雀 鯛 科 ) 和蝴蝶魚 ( 蝴蝶魚科 ) 等食草魚類,它們可以咬住藻類的碎片,而食草魚類如藍鰭 鯛 科和 鸚嘴魚 ( 鸚嘴魚科 ) ,它們會將藻類 從基質 上刮走。
以浮游植物為食的浮游動物被水體中的 鯡 魚屬 沙丁魚 ( clupeoides ) 和 雀鯛 魚類吃掉。珊瑚上的藻類被蝴蝶魚和 鸚嘴魚刮食 。一些碎屑被 鯔 魚吃,但碎屑無脊椎動物被各種食肉魚類所吃,包括蝴蝶魚、咕嚕魚和 鯛 魚。在食物網的頂端是食肉魚類。這些物種包括適合生活在珊瑚礁上的物種,其特徵是要麼像石斑魚那樣身體深 沉 ,要麼 像 煙管魚 一樣細長。攻擊方式包括埋伏躲避獵物和利用暗礁或其他魚類提供的掩護跟蹤獵物。 角叉魚 甚至被描述為迫使其細長的、磨蝕性的鼻子進入海鰻的喉嚨,並以鰻魚的胃內容物或腸道為食 (Johannes, 1981) 。在水體中的梭魚,當它們變得脆弱時會攻擊它們。
對於澳大利亞大堡礁上的 一棵樹礁 ,估算了由不同攝食組代表的礁上魚類總重量 ( 生物量 ) 的比例 (Goldman 和 Talbot, 1976 年 ) 。浮游生物占生物量的 10% ,包括珊瑚和 藻類食草動物 在內的食草魚類占生物量的 18% 。其餘食肉魚類占總數的 54% 。 表 6.3 顯示了太平洋和加勒比海珊瑚礁上七 個 食性 同功群的 物種相對比例。
溫帶的岩礁種類繁多。沒有珊瑚飼養員協會, 食草動物 物種在 40 ° 緯度以外也沒有太大的延伸。北部的 一個食草動物 例子是地中海 藍粘魚 ( blenius sanguinolentus ) 。雜食動物的典型例子是加利福尼亞的科蒂魚 ( clinocottusglobiceps ) ,它以藻類、海洋無脊椎動物和其他無脊椎動物為食。食 肉性礁魚以藤壺 和貽貝等 固著 無脊椎動物和活動無脊椎動物為食,包括腹足類、螃蟹、對蝦和其他甲殼類動物 (Gibson, 1969 、 1982 ; Grossman, 1986) 。
表 6.3 太平洋與加勒比海珊瑚礁上七 個 食性 同功群的 魚類物種百分比
6.5.2 大陸棚的攝食生態學
大陸棚靠近陸地和水域較淺,確保了它們的生產力。這些魚類的資源基礎是浮游植物的初級生產、生物的有機物和廢物以及陸地來源的有機物。在寒冷、營養豐富的水域上升的地方,如秘魯沿海,初級生產力特別高。
遠海區通常以 鯡魚科 沙丁魚 (clupeoid) 魚類及其捕食者為主。它們要麼直接以浮游植物為食,要麼以浮游動物為食,要麼兩者兼而有之。梭子魚和其他小型遠海魚類和大型無脊椎動物是包括鯊魚、 傑克魚 ( 鰺 科魚 ) 和鯖魚在內的遠海食肉動物的獵物。底棲魚類捕食正遊動在底部上方的獵物, 即底棲 動物獵物,或是生活在底層的魚類和大型無脊椎動物,或是捕食生活在底層的獵物 — 底棲動物。小型魚類,通常不到 60-80mm 長,以小型底棲動物為食,尤其是哈氏 橈足類 。較大的魚類,包括大型幼魚和成年 鰩 魚和比目魚,吃包括軟體動物和多毛類動物在內的大型動物群。 圖 6.4 給出了北海組合的簡化食物網,其中包括對每 個 飼養 同功群中 生物量的粗略估計 (Yang, 1982) 。
在西非洲幾內亞灣的大陸棚上,包括珊瑚礁和軟沉積物, Longhurst 和 Pauly(1987) 發現了幾種食性動物群落:食草魚類 (7 種 ) ;食浮游魚類 (11 種 ) ; 食礁表 有機物魚類 (13 種 ) ;底棲魚類 (18 種 ) ;活躍海底生物的食肉魚類 (6 種 ) ;在沿海水域捕食 活躍底棲生物 和魚類的食肉魚類 (12 種 ) ;在近海水域 活躍的 底棲魚類和魚類的食肉魚類 (17 種 ) ;以魚類、甲殼類動物和底棲動物為食的雜食魚類 (29 種 ) ; 遠海食肉 魚類 (34 種 ) 和 底棲食肉 魚類 (9 種 ) 。
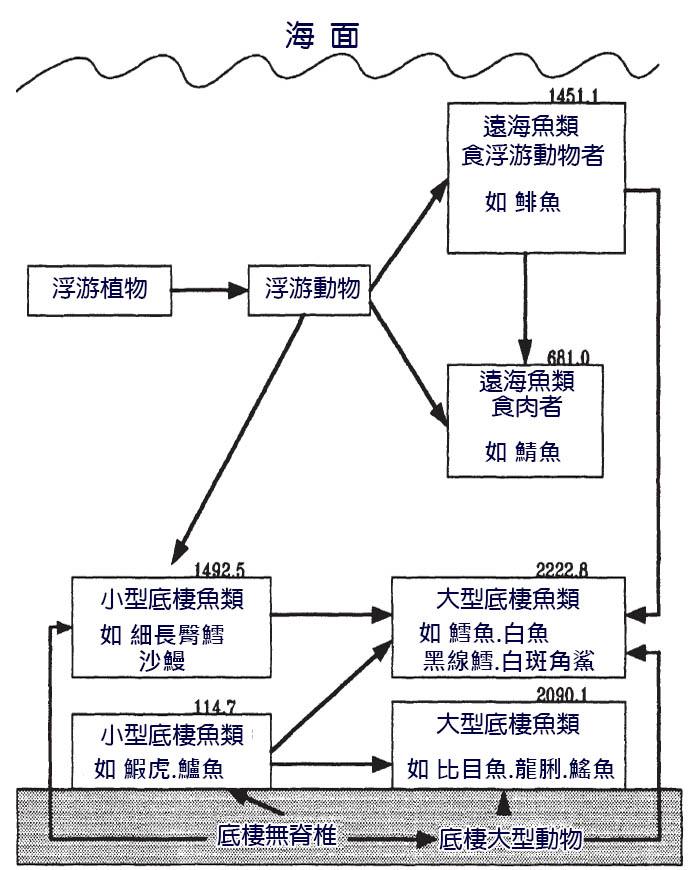
圖 6.4 北海的簡化食物網,包括攝食 同功群的 生物量估計值( x103 噸)。 Yang (1982).
6.5.3 遠海養殖業
由於環境的同質性和初級生產力的低,開放海域允許的謀生方式較少。主要的能量輸入是水體上部光照區浮游植物的初級生產。在能夠進行光合作用的區域下方,能量來源是從生產性海岸帶上方或側面輸入的有機物質。這些物質中的一部分通過水體下 沉 ,另一部分由沿著大陸棚向下流動的濁流攜帶,第三 個 來源是動物在生產力較高的上層水域和生產力較低的較深水域之間的活躍遷徙。
在海洋上層帶,大多數魚類都是一般的食肉魚類,通過追逐和抓取技術捕食較小的獵物。浮游動物包括一些生活在該區域內的物種,如飛魚和一些夜間從較深的中層帶遷徙覓食的魚類,白天返回深水區。 上層食 魚魚類,包括鮪魚和海豚魚,主要以遠海當地魚類為食,而不是以遷徙魚類為食。大鯊魚是最主要的食肉動物,包括鮪魚、較小的鯊魚和 鯨目 動物都是目標獵物 ( 圖 6.5) (Longhurst 和 Pauly, 1987 年 ) 。
在海洋中層帶,有三個餵食 同功群 (Marshall, 1979) 。小顎食肉動物以浮游動物為食,包括垂直遷移到上層帶覓食的物種;例如 小型角吻亞 目和燈籠魚 ( Myctophidae ) 。有遊動膀胱的大型捕食者包括深海星食魚 ( Astronethes . ) 。第三 個同功群由 沒有游泳囊的食肉魚類組成,其中包括一些類氣孔石首魚 ( stomiatoids ) 。
在深海區更深層和食物更貧瘠的地方,捕食者身體肌肉和骨骼減少,這可以解釋為對能量不足的環境的適應。典型的深海食肉動物是具有生物發光誘餌 的角魚 (Marshall, 1979) 。
相比之下,與海底相關的魚類、底棲動物和底棲物種擁有發達的骨骼和肌肉系統,這表明,即使在開闊的海洋下,水和沉積物之間的介面至少與上面的水域相比,資源豐富。有些物種是底棲食肉動物,捕食底棲動物和表殼動物。這類物種包括銀 鮫 、海 蜥魚和類鰻魚 ( 通氣魚科 ) 。其他物種從底層上方的水體中捕食獵物,可能是大型無脊椎動物或魚類。例如鼠尾 鱈 、鯊魚和深海鱈魚 ( Sedberry 和 Musick , 1978 ; Marshall, 1979) 。
遠海的魚類通常是多面手而不是專門的食性動物。這反映了在食物短缺的環境中,需要採取現有的食物。
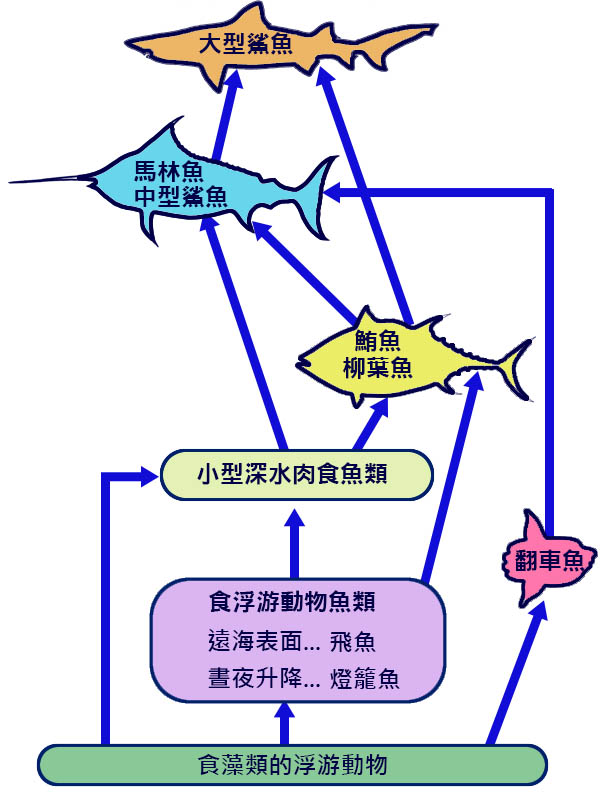
圖 6.5 熱帶、海洋上層系統的簡化食物網。 Longhurst and Pauly ( 1987) ..
6.5.4 南極魚類的攝食生態學
另一個極端環境的魚類,南極洲的海洋,也往往是通用餵養者 ( Everson 1984) 。這些海洋的一個不尋常的特點是,它們不包含任何密集的淺層、浮游的中上層魚類,如大多數其他海洋的 小鱗犬牙南極魚 。在南極周圍的海洋中,有 四種食 肉魚類:以魚和磷蝦為食的食肉動物,例如象鼻魚;浮游生物的食肉動物,例如,臭 鼬 ;大型遊動的無脊椎動物和生活在海底附近的魚類的食肉魚類,例如 Raja geogianus ;以及以底棲生物為食的食肉動物,包括多毛類、軟體動物和甲殼類動物,例如 長鰭金槍 魚 。一些南極物種也會吃大型藻類。
6.6 食物的檢測和選擇
對食物的檢測和選擇的研究大多集中在視覺捕獵食肉動物上,對其他感官或其他飲食的關注較少。
6.6.1 碎屑岩和食草動物的檢測與選擇
對於不流動的食物,檢測的問題可能並不嚴重。魚類面臨的主要問題是選擇。莫桑比克羅非魚 Sarotherondon mossambicus ,選擇性地以高品質的碎屑為食 (Bowen, 1984) 。魚用來選擇的線索是未知的。 食草性 魚類也是有選擇性的 (Horn, 1989) 。 珊瑚礁食草動物 的飲食並不反映藻類的比例。 這些食草動物 白天進食,因此視覺選擇不同顏色和形狀的植物可能會起到一定作用。藻類也可能會被排斥,因為它們的次級化合物使它們變得不好吃。藻類的質地也可能與魚類避開難消化的植物有關。
6.6.2 食肉動物的檢測與選擇
許多食肉魚類通過視覺發現獵物,而獵物的視覺特徵也在決定獵物是否被吃掉或拒絕的過程中起著重要作用。實驗研究表明,獵物的重要視覺特徵與背景、大小、運動、形狀以及某些情況下的色調形成對比。
魚能探測到潛在獵物的最大距離就是反應距離。這取決於獵物的大小 ( 對於較大的獵物,距離更大 ) 、水的濁度和魚眼的分辨力。在水體中遊動的魚可以被認為是在視覺上搜索圓柱體,圓柱體的尺寸由反應距離 和魚遊的 速率決定 (Eggers, 1977) 。幼魚由於遊動速度慢,眼睛分辨力差,只能搜索到很小的體積。沙丁魚幼體 ( clupeoids ) 每小時的搜索量可能低於 1 公升 ( Blaxter 和 Hunter, 1982 年 ) 。如果存在多個獵物,那麼對魚來說最大的獵物就是可能被攻擊的獵物 (Brien 1976) 。獵物 的表觀大小 取決於它的絕對大小以及它與魚的距離。
使用不同獵物類型的研究有助於確定魚類視覺上選擇獵物的線索。三刺 棘背魚 對紅色、快速移動、又長又窄、體型較大的獵物更為偏愛 ( 其中,大型與捕食者的嘴大小和獵物的大小範圍有關 )(Ibrahim 和 Huntingford , 1989 年 ) 。這些偏好並不是絕對的,而且 粘背魚 是 不 挑食的餵食者,其飲食在很大程度上受限於它自身的小尺寸 ( 長度不到 80mm) 。
在夜間,或在混濁的水中或對於距離大於視野的獵物來說,其他感官資訊將變得重要。人們對非視覺餵養的瞭解要少得多。有些魚可能通過聽覺察覺獵物。鯊魚可能會被游泳者所發出的聲音所吸引,因為這種聲音類似於一條大而受損的魚發出的聲音 ( Budker , 1971) 。獵物的運動也會產生低頻壓力波,由側線系統探測到。失明的深水 甲杜父魚 Cottus bairdi 能有效地捕食移動的獵物,但如果側線系統被阻斷,這種反應就會被抑制 (Hoekstra 和 Janssen, 1985) 。有些魚能探測到活獵物產生的電場。鯊魚和 鰩魚利 用它們的電感受器,壺腹器官,可以找到包括埋藏在基底中的比目魚在內的獵物 ( Bleckmann , 1986) 。一些硬骨魚,特別是 裸臀魚科 、裸 背刀魚科 和 大象魚科 ,可以通過魚產生的微弱電場產生的扭曲來探測獵物 ( Bleckmann , 1986) 。 ( 此外, 南美洲的電鰻可以通過其電器官的高電壓放電電擊 暈 獵物。 ) 許多以底棲獵物為食的物種 ,在觸鬚或鰭鰩等附屬物上都有化學感受器,可能用它們來探測底層或其下的獵物。在三角魚科中,胸鰭的底部射線與鰭分離,用作觸角,在海底尋找食物。
從委內瑞拉奧里諾科河下游主河道採集的一組裸體動物完成了捕捉獵物的非凡壯舉 (Lundberg 等人 , 1987 年 ) 。以浮游生物為食。它們不知何故在快速流動的混濁、變色的水中發現並捕獲不到 1mm 長的獵物。由於缺乏光線穿透力和獵物體型較小,獵物的視覺和化學反應都不太可能發生。
6.6.3 最佳覓食
食物可以根據它們對魚的價值來選擇。一個理論體系,即最優覓食理論,已經被發展用來預測動物的飲食結構,前提是覓食動物能毫無阻礙地獲得各種食物 (Stephens 和 Krebs, 1986) 。換言之,選擇食物是為了使覓食者的利潤最大化。衡量利潤的最佳選擇通常假定為淨能量收益率,即 採 食者單位時間內獲得的總能量減去覓食的能源成本。這顯然取決於每種獵物的能量含量,遇到特定類型獵物所需的時間,以及搜索和捕獲該獵物類型的能量成本。在它的經典形式中,這個理論做出了簡單而明確的預測 (Hart, 1986) 。當捕食者連續和隨機地遇到獵物時,應按獲益能力的降低標準選擇獵物,直到包括一個獲利能力較低的獵物導致總體淨能量增益下降。飲食選擇只取決於更有利可圖的方案的密度,而不取決於那些在飲食中沒有獲益的方案的密度。
從品質上講,魚可以以一種接近理論預測的方式覓食;然而,在數量 上,魚並沒有達到所預測的、最佳的飲食結構。這些魚繼續在它們的飲食中包含獲益較低的獵物,即使在更有利可圖的獵物密度達到了可以忽略前者的水準之後。以 不同大小的水蚤為食的藍鰓太陽魚,隨著大型獵物密度的增加,它們的飲食中包含了更多更大、更有利可圖的水蚤,但也繼續吃 一 些小的,利潤較低的水蚤 (Werner 和 Hall, 1974) 。然而,最佳覓食理論的使用 使沃納 和他的同事成功地預測了湖泊中 藍鰓類動物 捕食的獵物大小譜的季節性變化 (Werner 和 Mittelbach , 1981) 。必須修改優化覓食理論的簡單形式,以考慮到覓食者動機變化的影響、覓食者覓食時的風險及其行為能力 (Stephens 和 Krebs, 1986) 。
6.7 攝食生態形態
6.7.1 篩檢程式 饋 線
七 鰓鰻的梭形 幼體在淡水中的軟沉積物中穴居。它以藻類如苔蘚、矽藻和碎屑為食。幼體 通過鰓籃的 交替收縮和擴張來維持水在過濾裝置上的單向流動。水從口腔進入咽部,通過一個 支鏈狀 的網路,其中排除了任何大顆粒。較小的顆粒被抓取在 鰓和咽壁 分泌的粘液中。水流很慢,很高比例的顆粒會被截留, (Hardisty and Potter, 1971) 。
在濾食性鰓類 和其他硬骨魚類中,過濾通常發生 在鰓弓的 突出物上,鰓 耙又細又密 。在沙地 鯡 魚 ( Dorosoma cepedianum ) 中,不同大小顆粒從水中過濾的有效性 與耙距有關 ( Mummert 和 Drenner , 1986) 。隨著魚的生長,距離趨於增加,因此較大的魚在過濾較小的顆粒方面是不太有效的。隨著沙地 鯡 魚的生長,浮游植物在它們的飲食中變得不那麼重要了。 姥 鯊 和 鯨鯊 也會用鰓耙過濾浮游動物。那些 姥鯊 很 可能是在冬天開始的時候脫落的,那時鯊魚進入了更深的水域 (Stevens, 1987)) 。
6.7.2 腐食
耙鰓也可能 在碎屑岩中的食物分類中發揮作用。 鯔 魚對口腔中的沉積物進行分類,丟棄無機物,同時保留耙 耙 上的有機碎屑 (Bowen, 1984) 。一些碎屑有肌肉 磨胃。 在有害的慈 鯛 科魚類中,胃是薄壁的,但能夠在胃液中產生極低的 pH 值。碎屑動物的腸道通常 比食草 動物或食肉動物的腸表面積大。這可以通過一個 長腸來實現 — 在鯉魚類的 陰唇中,腸是 體長的 15-21 倍 — 或者是通過有發達的粘膜皺 襞 來實現的。 參圖 6.6 。
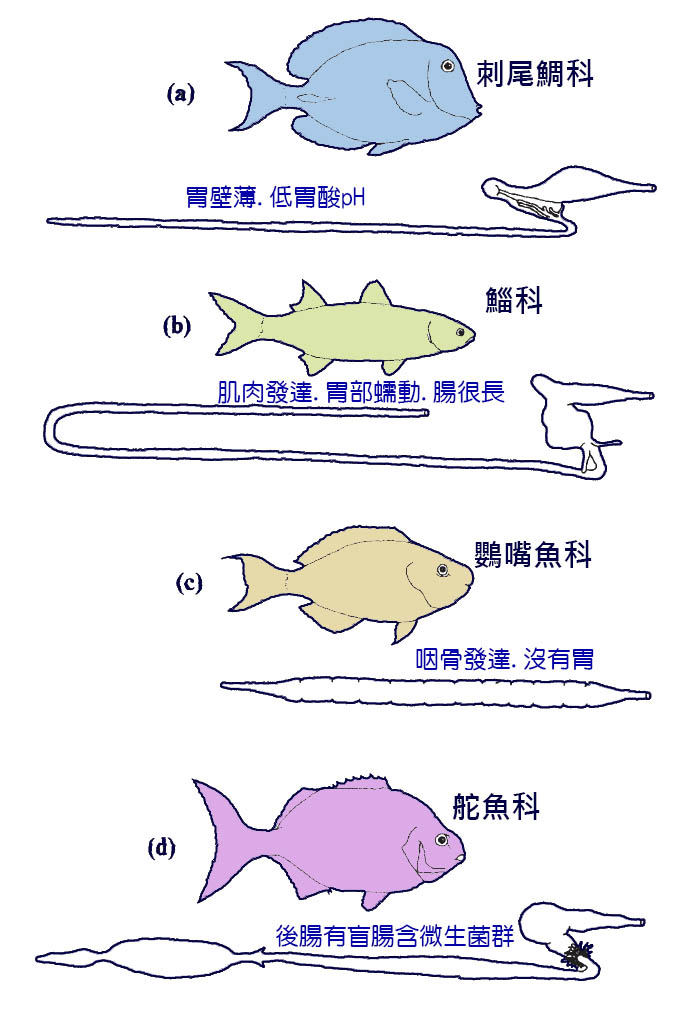
圖 6.6 草食魚類的腸道適應:( a )薄壁胃,低胃酸 pH ,如刺尾 輖 ;( b )肌肉發達、胃部蠕動、 腸很長 ,如 鯔 魚;( c )發達的 咽骨,無胃,如鸚嘴魚 ;( d ) 後腸含盲腸 的微生物菌, 如舵魚 。 Horn (1989).
6.7.3 食草魚類
食草魚類和 食肉 魚類都有一個問題,那就是他們的食物很難消化。為了獲得有用的營養,草食性魚類必須打破植物細胞壁。有四種解決問題的方法 ( 圖 6.6)(Horn, 1989 年 ) 。在包括 食草性 麗 魚科 和 某些 刺尾魚 ( 刺尾 鯛 科 ) 的群體中,胃壁薄,產生高酸性胃液。 腸很長 。 顎齒的 形狀使魚能夠修剪植物材料。在其他水 豚 魚和 鯔 魚中,胃是厚的 ・ 嚎叫,並充當砂袋,其中的物質是粉碎。鸚鵡魚的牙齒融合在一起形成一個像鸚鵡一樣的喙。有一個高度發達的磨坊,當食物 通過咽時 ,食物被碾碎在 咽骨之間, 儘管沒有胃, 但腸是 適度長的。第四種是不常見的,但在南半球以褐色海藻為食的脊椎動物中被發現。這些魚有一個 長腸, 在它們的後端有一個後腸盲腸。它有一個血管化良好的細胞壁,裡面有一個微生物群落。據推測,微生物有助於分解植物物質並產生 可被魚吸收 的物質。這個解決方案類似於反芻哺乳動物使用的機制。
長的 腸道使魚 能夠處理大量的食物。在某些物種中,食物快速通過腸道,魚通過快速加工大量劣質食物獲得營養和能量 (Pandian 和 Vivekanandan , 1985) 。對於那些依賴共生微生物來幫助它們消化頑固不化的植物物質的物種來說,情況並非如此。
6.7.4 食 浮游物魚類
當食浮游 物魚類體型較小且密度較高時,一些浮游動物的魚類會過濾飼料。它們 張開嘴遊過 獵物聚集的地方,獵物被收集在 細而密的鰓耙上 。在實驗室實驗中,北鳳尾魚 以鹵蟲 (Artemia) naufile 為食 ( Blaxter 和 Hunter, 1982) 。對於較大的獵物,魚通過撞擊單 個 顆粒來進食。浮游動物的口通常很小,呈管狀。在某些物種中, 顎是突出 的,魚抓起獵物就好像用吸管收集獵物一樣。浮游動物的腸道通常 比食草 動物或碎屑動物的腸道短。如果 存在咽骨 ,則其牙齒細小且間隔緊密 ( 圖 6.2) 。
6.7.5 食軟體動物魚類
雖然蝸牛的肉品質很好,但必須從 保護殼 中提取出來。有些魚通過抓起暴露在外的腳和頭,或者在軟體動物將它們安全地收回之前抓住虹吸管,從而達到這一目的。其他物種有特殊的形態使它們能夠粉碎貝殼。在底層居住的滑板 魟 魚中, 顎齒被 壓平,形成一個壓碎的路面。在許多軟體動物中,貝殼被壓碎在 咽骨之間,咽骨 上長著低矮的圓形壓碎牙齒 ( 圖 6.2) (Fryer 和 Isles, 1972) 。食軟體動物魚類南瓜子太陽魚不同於浮游動物的藍鰓太陽魚,它的 咽齒寬而鈍, 而不是 細而密的 ( Keast , 1978) 。
6.7.6 食肉魚類
大多數魚類都是 食肉性的 ,在捕捉活生生且經常高度活動的獵物方面有著廣泛的解決方案。這裡只提到幾個。
成年的七 鰓鰻沒有 下顎,但仍然是一種可怕的捕食者 (Hardisty and Potter, 1971) 。七 鰓鰻附著 在它的獵物上,通常是一條骨瘦如柴的魚, 它的口盤充當 吸盤 ( 圖 1.5) 。一旦附著,七 鰓鰻用 舌頭上的牙齒 刺入肉中 。然後,獵物的體液和破碎的組織被 泵入咽 部。 魚盤和 舌頭產生的吸力阻止獵物將附著的七 鰓鰻趕走 。
雪茄 達摩鯊 ( Isitius brasiliensis ) 的嘴唇可以充當吸盤。鯊魚依附在一條較大 的魚上 ,然後用它像手術刀一樣的牙齒切出 一個肉塞 ( Stevens, 1987 ) 。生活在 最 底層的 絞口鯊 ( Ginglymostoma ) ,用吸力把獵物從洞裡抽出來。鯊魚用厚嘴唇做一個海豹,然後擴張 它的咽腔 ,產生吸力。
吸力是硬骨魚的基本進食機制。當口腔擴張後,魚突然張開嘴, 將水吸進去 ,並隨之捕食。這種進食方式在大嘴食肉動物 ( 如雙體魚和 垂釣者魚 ) 中最為發達。
捕食一系列獵物的食肉動物往往具有普遍的形態特徵。下頜和咽牙齒,當存在時,是簡單的錐體;還有肌肉發達的胃和相對較短的腸。在 胃和腸的 交界處可能有 一系列盲端囊 ,即 幽門盲囊 ,但這些囊也存在於 一些食草動物 和有害生物中。鮭魚和鱒魚 ( 鮭魚科 ) 和 鱸魚 ( 鱸魚科 ) 是這種多面手食肉動物的好例子。
許多食魚魚類在形態上表現出 一種趨同現象 ,這種形態可以用梭子魚來描述 ( 圖 3.10) 。身體被拉長,其長度上的深度大致相同。鼻子很長,有點扁平,下顎上有許多鋒利 的後尖牙 。除了梭子魚和它的近親 ( 魟 科 ) ,其他表現出這些特徵的魚類還有一些食魚的 麗 魚科 和 甲 梭魚 (( 金梭魚科 Sphynaenidae ) 科 ) 。
在水生食肉動物的頂端是大鯊魚,如大白鯊 (Carcharodon carcharias) 。這種和類似物種的下顎上有一個鋸齒狀邊緣的令人生畏的三角形牙齒。軟骨魚類動物的牙齒 來源於板狀 鱗片,並在一生中不斷更換。它的嘴很大, 胃很大 ,這樣大的獵物就可以被切開、吞咽和消化。最大的鯊魚捕食其他食肉動物,包括鮪魚、中型鯊魚和海洋哺乳動物,包括海豹和海豚。
6.7.7 生態形態學假說
這個 假設假設假設 形態與生活方式密切相關,因此可以預測生活方式。第三章描述了體型與棲息地的關係。根據這個假設,飲食應該 可以從魚的 形態來預測,特別是從與攝食有關的形態特徵, 如嘴的 大小, 顎的形狀 和牙齒的牙齒。該假說還預測,形態相似的物種有相似的飲食結構。
莫塔 Motta (1988) 對這一假說進行了優雅的研究,他比較了 十種礁棲蝶魚 的形態和飲食。這些物種包括一個浮游動物,珊瑚上的流覽器,一個流覽雜食動物和一個海底雜食動物。口腔和 頜骨 形態的變化範圍是驚人的 ( 圖 6.7) 。莫塔 Motta 的結論是,形態不一定是一個良好的飲食預測,但是一個良好的預測如何魚類飼料。餵食機制是決定飲食的一個重要因素。即使是形態特殊的物種,在浮游動物數量豐富時,也會從通常的食物轉向捕食浮游動物。
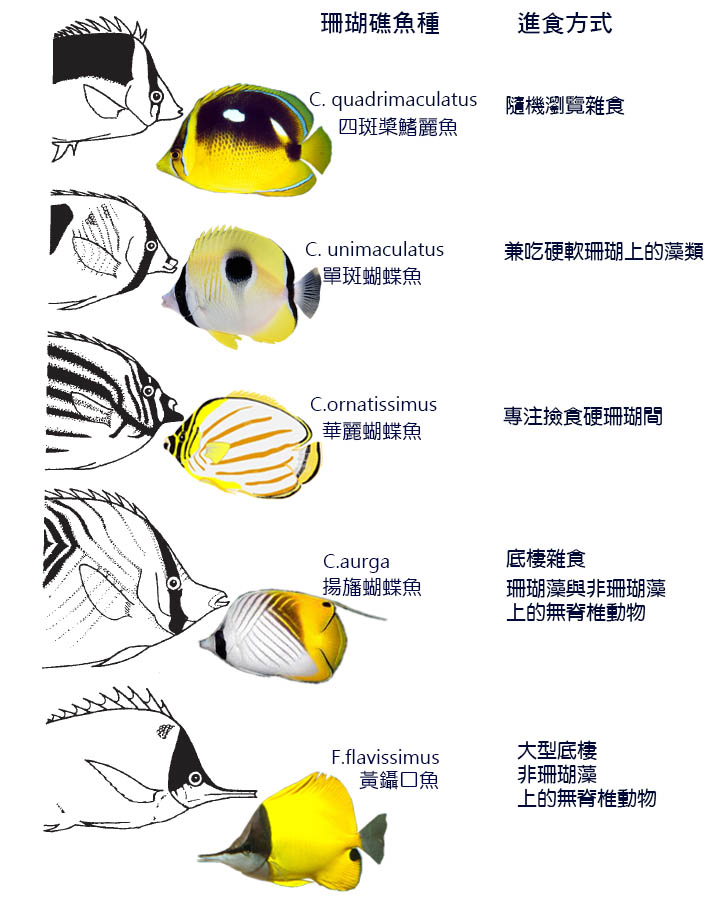
圖 6.7 蝶魚嘴 的大小和位置與日常飲食的關係。 Motta (1989).
6.8 魚類營養種類
由於魚類攝食行為的幾個特點,將魚類劃分為明確的營養類別通常很 困難。許多物種在其一生中都表現出飲食的重大變化,即個體發生的變化。這些變化通常與魚的體型隨著年齡的增長而增加有關,這使得魚可以捕食更大的獵物。個 體發生的變化會突然發生。以魚為食的梭子魚和大口鱸魚在生命早期就從無脊椎動物轉變成了魚類,這種偏好在他們的餘生中一直保留著。相比之下,鱸魚的飲食變 化較慢,隨著體型的增加,逐漸變得更加愛吃魚。在一些種群中,鱸魚可能永遠達不到魚成為食物重要組成部分的大小。在其他物種中,個體發生的變化與食物聚集 的方式有關。在一些 鯡魚科 沙丁魚 ( clupeoids) ,包括秘魯的 鯷 魚 ( anchoveta) ,年輕的主要是浮游動物。隨著魚的生長, 鰓耙的間隔 變小,成年鳳尾魚的飲食中包含了相當一部分的浮游植物。
許多物種都是多面手的食性動物,所以它們的飲食會隨著食物供應的季節性變化而變化,或者隨著魚進入不同的棲息地,甚至是因為突然出現了大量的適宜食物。瞭解和預測魚類營養行為的這些變化將是全面瞭解魚類在水生群落中的作用的關鍵之 一 。
6.9 食物利用
利用食物的原則對所有魚類都是共同的,不管它們生活在哪裡和如何生活。
6.9.1 能源收支
食物中的能量要麼在新陳代謝過程中被用來做有益的工作,包括組織的修復和維護、食物加工和游泳,要麼作為積累的組織的化學能儲存起來。熱力學定律意味著能量的輸入必須等於輸出,當能量被用來做有用 的功時 ,一些能量會以熱量的形式散失 ( Brafield 和 Llywellyn , 1982) 。這可以用一個簡單的方程式來表示:
A=R+P
其中 A 是被同化食物的能量, R 是新陳代謝過程中損失的熱量, P 是魚以生長的形式獲得的能量。被同化食物中的能量, A ,比所消耗的食物中的能量 C 要少,因為這些能量中有一部分在排泄的糞便 (F) 中損失,而另一部分在蛋白質 (U) 的新陳代謝過程中產生的含氮排泄物中損失。因此,以下方程式描述了能源預算:
A=C-(F+U)
A=R+P
式中 (A/C) 是對消耗的食物被同化的效率的衡量。就所消耗食物的重量而言,食肉動物的效率 高於食草動物 或碎屑動物,因為 後者攝 入的消化物質相對較多。 (P/C) 是衡量消耗的食物轉化為增長的效率的一個指標,它是總增長效率。生長效率也可以用同化食物來表示 (P/A) 。
對食肉性 和草食性魚類的能量預算進行了彙編,使得 Brett 和 Groves(1979) 能夠繪製出平均能量預算,從而粗略地描述了食物中的能量是如何分配的。食肉動物的平均值是:
100C=44R+29P+27E
對於食草動物 :
100C=37R+20P+43E
式中 E=(F+U) 。
這些數字表明,作為一個食肉魚類,它的能量消耗稍微高一些 (44R 相對 37R) ,但是食肉魚類更有效地吸收食物 (27E 相對 43E) 。
6.9.2 營養素收支
食物提供營養和能量,雖然食物的成分可以分解,合成或消除新的化合物,但氮、 磷和其他 元素的預算必須用平衡方程來描述:
收入 = 留存 + 消除
殘留部分是魚體中所含的量。被排除的部分會流失給魚,但魚的排泄物作為營養來源對群落中的其他動植物可能很重要。珊瑚礁魚類以成片的海草或紅樹林沼澤中的珊瑚礁為食,將 氮和磷 等營養物質輸入珊瑚礁系統,以糞便和氨水等可溶排泄物的形式釋放出來。
對於生活在波蘭低地溪流中的一組物種, Penczak (1985) 估計,每保留一公斤元素,魚就要消耗大約 8.9 公斤碳、 7.6 公斤氮和 4.1 公斤磷。在魚 的乾重中 , 碳約占 42% , 氮約占 10% , 磷僅略 高於 2% 。這些營養物質被鎖在魚的 生物量裡 。
6.10 食物消耗率
正如能源和元素預算所表明的,可用於活動的能源和材料 R 和增長 P 將取決於以食物形式存在的收入率。食物的消耗率將取決於環境中食物的可獲得性和魚的進食準備情況。
功能反應曲線描述了魚類所攝取食物的數量與其在環境中的豐度之間的關係。這通常是漸進的 ( 圖 6.8) 。有一個食物密度,超過了這個密度,攝食率就沒有進一步的提高,魚正在以最大的速度進食。
這個最大速率又與魚的大小和環境溫度有關。 當魚越長越大 ,它對食物的絕對需求就會增加,但是在許多物種中,單位重量的食物消耗量會下降。相對而言,隨著魚變大,它們的消耗量也會減少。這可以用一個簡單的模型來描述,其中 Cmax 是最大消耗率, W 是魚的重量:
Cmax = nW m
m 通常取小於 1.0 的值,表明這種關係 是異速異構 的 ( 圖 6.9) 。
在魚可以忍受的溫度範圍內,消耗率首先增加到最大值,但溫度的進一步升高會導致消耗量下降 ( 圖 3.2b) 。其他因素,如鹽度和 pH 值也可能影響消耗率,但其影響研究較少。在一項關於 褐 鱒魚的實驗研究中 , 10 ℃ 下的每日最大消費量估計值為 6.3% 體重 (5 克體重 ) 和 2% 體重 (300 克魚 )(Elliott, 1975) 。 15 ℃ 時的當量範圍為 11.3% 至 5.3% 。對各種自然種群的觀察表明,對許多物種來說, 0.5-5% 的體重消耗率是典型的。
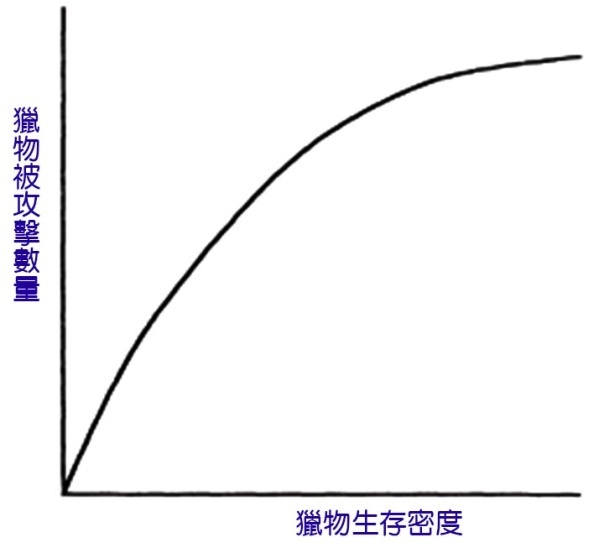
圖 6.8 理想功能反應曲線顯示了獵物密度和被攻擊數量之間的漸近關係。
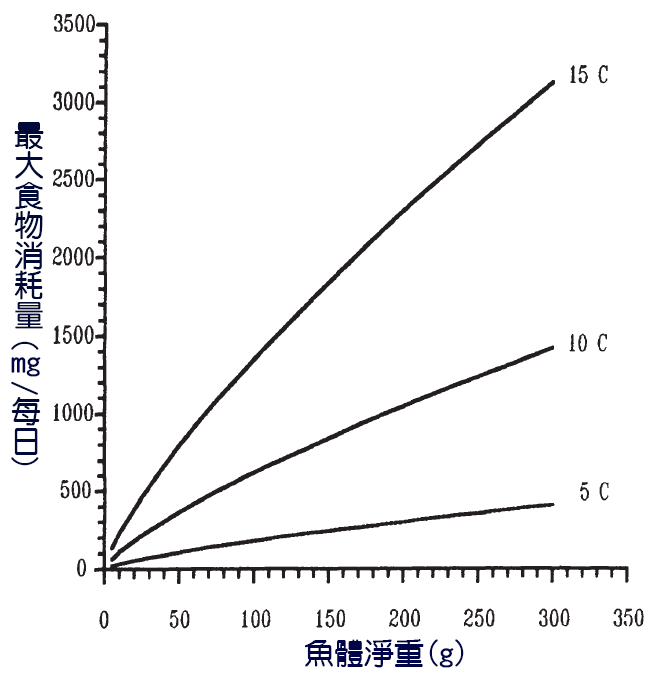
圖 6.9 在 -5 ℃ 、 0 ℃ 和 15 ℃ 三種溫度下褐 鱒 ( Salmo trutta )的體重與最大每日攝食量的關係( C= 攝氏溫度)。計算的曲線。 Elliott (1975).
6.11 增長
與鳥類和哺乳動物不同的是,大多數魚類在性成熟後仍會繼續增加長度和重量,儘管隨著魚變大,生長速度會下降。觀察到的生長模式是由魚類基因型定義的生長潛力與魚類所經歷的環境條件相互作用的結果 ( 圖 1.7a) 。食物供應和溫度通常是最重要的環境特徵。 Weatherley 和 Gill(1987) 提供了魚類生長生物學的全面綜述。
6.11.1 魚類體型與棲息地的關係
魚類成魚的大小範圍很廣。最小的是熱帶 鰕 虎 大約是 10mm( Miller, 1979) ) ,最大的是鯨鯊,它的長度可以達到 15 米或更多。這些差異是由生長模式的差異造成的,即使是在一個物種 內部, 這種差異也會有很大的差異。
在一個地區的全部 魚類區系中 ,在淡水、淺水和深水 (>2000 m) 海洋中,典型成熟尺寸小於 100 mm 的魚類比例從熱帶到極圈區逐漸減少 (Lindsey, 1966) 。 Marshall (1979) 描述了海洋中上層帶中小型魚類的重要性,特別是小型燈籠魚和氣孔魚。然而, Eversen (1984) 指出,許多南極特有物種也相對較小。 Miller(1979) 討論了有利於魚類小體型的生態因素。一個明顯的優勢是能夠在絕對食物量很少的環境中達到性成熟。能夠開發結構複雜的環境,如珊瑚礁或岩礁也是一個優勢。然而,所觀察到的全球格局的原因仍不清楚。
6.11.2 食物和其他環境特性對生長的影響
魚的生長依賴於魚獲得足夠的食物,但隨著魚的生長,它通常可以捕食更廣泛的獵物。這些飲食的改變會對魚的進一步生長產生影響。在一些多面手的食肉動物中,如鱸魚和鱒魚,隨著魚的尺寸的增長而轉向食魚,通常伴隨著生長速度的加快。這是因為體型較大的獵物利潤更高 ( Mittelbach , 1983) 。
魚類的生長率和消耗率之間的關係通常是 一條負 加速曲線 ( 圖 6.10) 。為了保持一個 恒 定的體型,需要最少的食物量,即 Cmain 。增長率與食物消費之間關係的一個結果是,在低於最大消費率 Cmax 的 日糧 ( Copt) 下達到最大生長效率。魚類不能同時使其生長速度和生長效率最大化 (Brett, 1979) 。
增長率還取決於食品的盈利能力 (Elliott, 1979) 。有一個食物顆粒的大小範圍,如果食物顆粒小於或大於這個最佳範圍,則允許最高的增長率,而這個增長率會下降 ( Wankowski 和 Thorpe, 1979) 。生長速率也隨溫度而變化。在最佳溫度範圍內,無 限制日糧餵養 的魚類生長速度最高 (Elliott, 1979) 。 對溫性狹窄 的褐 鱒 來說, 最 適生長溫度為 12-13 ℃ 。
生長還受到其他非生物因素的影響,包括氧氣和鹽分以及生物因素 (Brett, 1979) 。提供最佳生長條件的非生物和生物因數隨物種的生活方式而變化,並反映了它們的生活方式。其他魚類的存在可以通過干擾或開發競爭降低生長速度。在養魚場魚群密集的地方,魚通常必須定期按大小進行分級,以區分較大的優勢魚和較小的從屬魚。在 一些淺 水魚類中,同種魚的存在可能會提高生長速度。
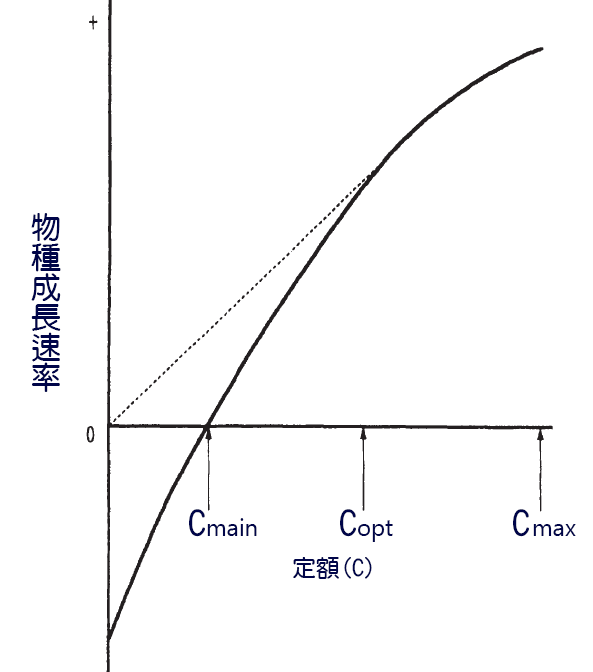
圖 6.10 每日進食與生長比率關係示意圖。虛線表示與原點相切,以確定最佳比率。
C main= 維持比率; C opt= 最佳比率, C max= 最大比率。
6.11.3 重量與長度的關係
到目前為止,人們主要從體重 ( 或能量含量 ) 的角度來討論魚的生長,但是魚的長度和體積都在增長 ( Weatherley 和 Gill, 1987) 。對於 給定種群 的魚類,其重量 W 和長度 L 之間的關係通常可以用以下等式來描述:
W= aL b
其中 b 是 3.0 ,如果魚是等距生長 ( 不改變形狀 ) 。如果魚在生長過程中改變形狀 ( 異速生長 ) , b 將與 3.0 不同 ( 圖 6.11) 。如果魚 變薄了 , b 小於 3.0 ,如果它變得更豐滿, b 大於 3.0 。因此,重量與長度之間的關係式給出了魚類種群狀況的一些指示。一種常用的 ( 如果是粗糙的 ) 狀態測量值 CF 是:
CF=W/L 3 。
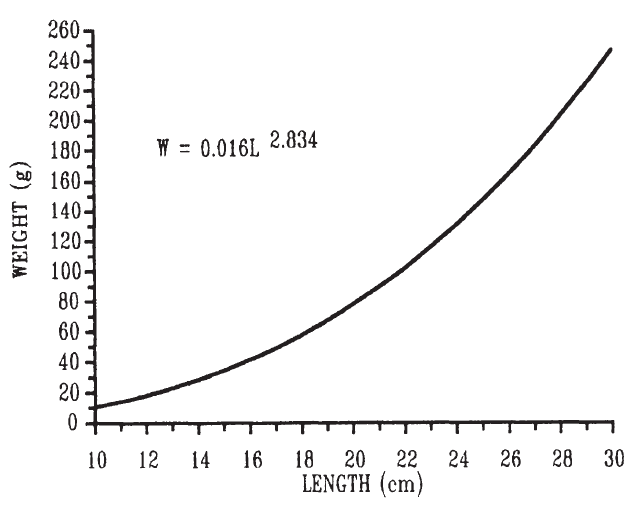
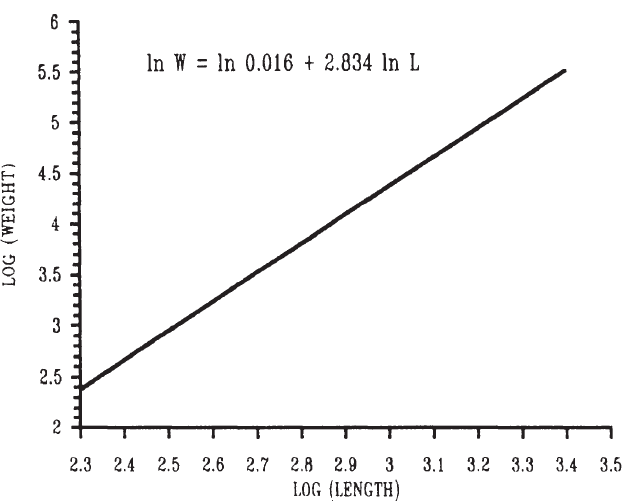
圖 6.11 褐 鱒 ( Salrna trutta)體重和長度之間的關係。 ( 左圖,數學數值;右圖,對數數值 ) Elliott1975.
6.11.4 生長測量
生長可以通過魚的絕對重量 ( 或能量含量 ) 或長度隨時間的變化來衡量。如果以足夠長的時間間隔測量魚的大小,以掩蓋季節或繁殖週期引起的短期尺寸變化,通常的生長模式是不對稱的 S 形。在漁業生物學中,魚一生中的生長模式通常用 von Bertallanfy 曲線描述,其形式如下:
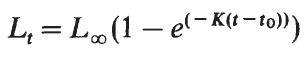
式中 , Lt 是魚在 t 年齡時的長度, L ∞ 是魚的漸近長度, t 是魚的長度為零的假設時間, K 是生長曲線接近漸近線的速率。雖然最初是從生理學原理推導出來的,但該方程被認為是對生長曲線的一個有用的數學總結 ( Beverton and Holt, 1957 ; Ricker, 1979) 。示例如 圖 6.12 所示。 von- Bertallanfy 曲線不太適合描述魚類的早期生長或連續測量生長的時間間隔很小。
絕對增長率是魚的大小的函數。體重從 1 克增加到 1001 克的魚,其絕對增長率與同一時期體重從 1 克增加到 2 克的魚相同,但後者的體重增加了一倍。與規模無關的增長率衡量指標是具體增長率,具體增長率可通過以下方式衡量:
(loge Sf -loge So)/t
式中 , Sf 和 So 是魚在指定時間段內的最終和初始大小, t 是時間段的長度。在魚的一生中,特定生長率在早期生命史階段通常很高,隨著魚的年齡和體型的增大而降低。如果一條魚沒有生長,它的特定生長率為零。
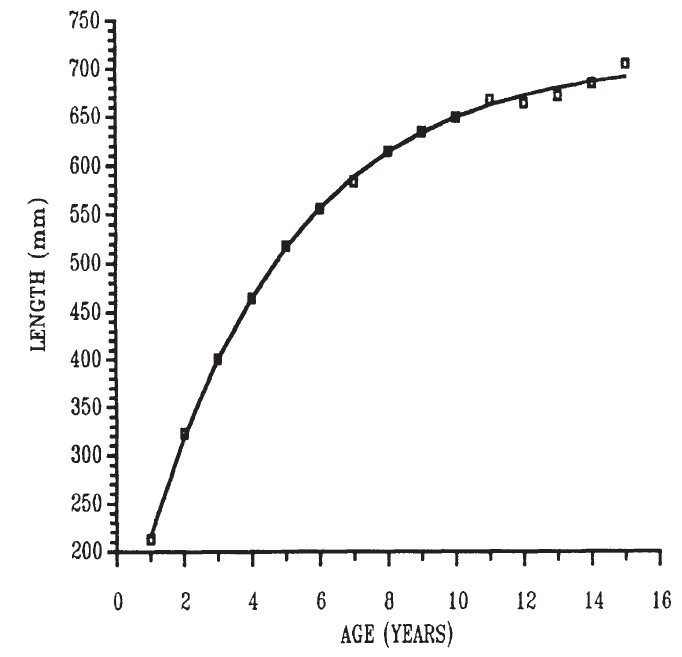
圖 6.12 安大略湖玻璃梭鱸長度的增長。 von- Bertallanfy 增長曲線已被擬合到數據中。曲線參數: L ∞ =712 mm ; K=0.0232 ; t o=-0.566 年。
6.11.5 魚的老化
在實驗室裡,魚的生長可以在一個已知的時間段內進行測量。然而,在自然種群中,生長模式通常需要根據已知年齡的魚類大小來確定。幸運的是,許多魚類的年齡可以確定,因為鈣鹽 沉 積模式的變化,如鱗片、內耳 耳 石、鰭 棘 、脊椎骨或其他骨骼 ( Summerfelt and Hall, 1987) 。
在硬骨魚鱗片中,隨著鱗片的生長,一系列同心 的脊狀物 或環狀物形成 ( 圖 7.13) 。這種 迴 圈模式的變化表明魚類生長速度的季節性變化或產卵等事件。像一系列緊密相連且每年重複出現的環狀結構稱為環狀。魚的年齡是通過 計算環空來 評估的。
內耳 的耳石可以 提供兩種類型的資訊 ( 圖6 .13) 。不透明和更透明的材料交替出現,標誌著生長速度的季節性變化,這些計數可以用來計算魚的年齡。在一個更精細的尺度上, 耳石物質 有一個每天的 沉 積模式。在幼魚中,這些 日環數可以 用來估計魚的年齡 ( 以天計 ) 。這項技術有望在解釋硬骨魚類最早生活史階段的生態學方面極有價值,並已被用來 確定礁魚在 其遠海階段結束時定居的年齡。
像 Agnithans 無 頜魚綱和 Elambobranches 板鰓亞綱 ( 軟骨鯊魚 ) 這樣缺乏骨質結構的魚類可能仍然會衰老。在七 鰓鰻中,類似耳石 的結構顯示出每年的帶狀分佈 (Beamish 和 Medland , 1988) 。在軟骨魚類動物中,椎骨或脊椎的部分可能顯示環狀。另一種不同的方法是利用長度的頻率分佈。如果從具有季節性繁殖模式的種群中測量大樣本的長度,則種群中存在的每 個 年齡級應顯示為長度 ・ 頻率分佈的峰值 ( 或模式 ) 。 一個年齡組將與前一個較年輕的年齡組和下一個年齡組分開,其長度相當於連續繁殖季節之間的長度 (Pauly 和 Morgan, 1987 年 ) 。
必須注意驗證對某一人群常規使用的老齡化方法。這可以通過比較幾種技術的老化結果和對控制年或日模式形成的因素的實驗室研究來實現。
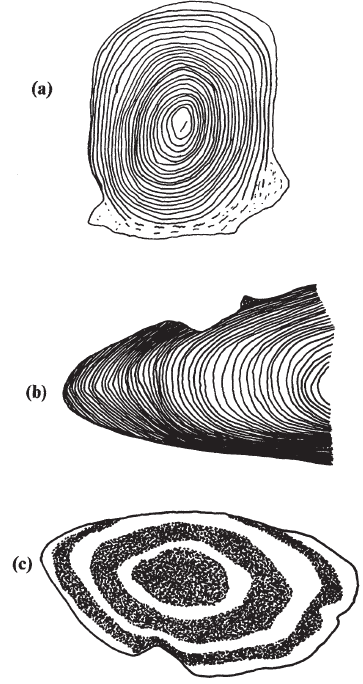
圖 6.13 硬骨魚類的年齡結構:( a )鮭魚的鱗片顯示年輪;( b )慈 鯛 幼 魚耳石 顯示年輪;
(c) 比目魚耳石顯示 年輪。